Department of Mechanistic Cell Biology, Max-Planck Institute of Molecular Physiology, Dortmund, Germany.
Department of Chemistry and Biochemistry, University of Colorado at Boulder, Boulder, United States.
Elife. 2017 Dec 27;6:e33442. doi: 10.7554/eLife.33442.
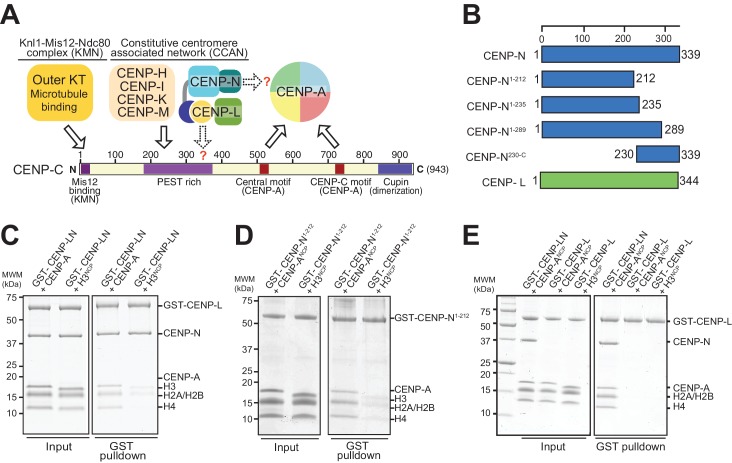
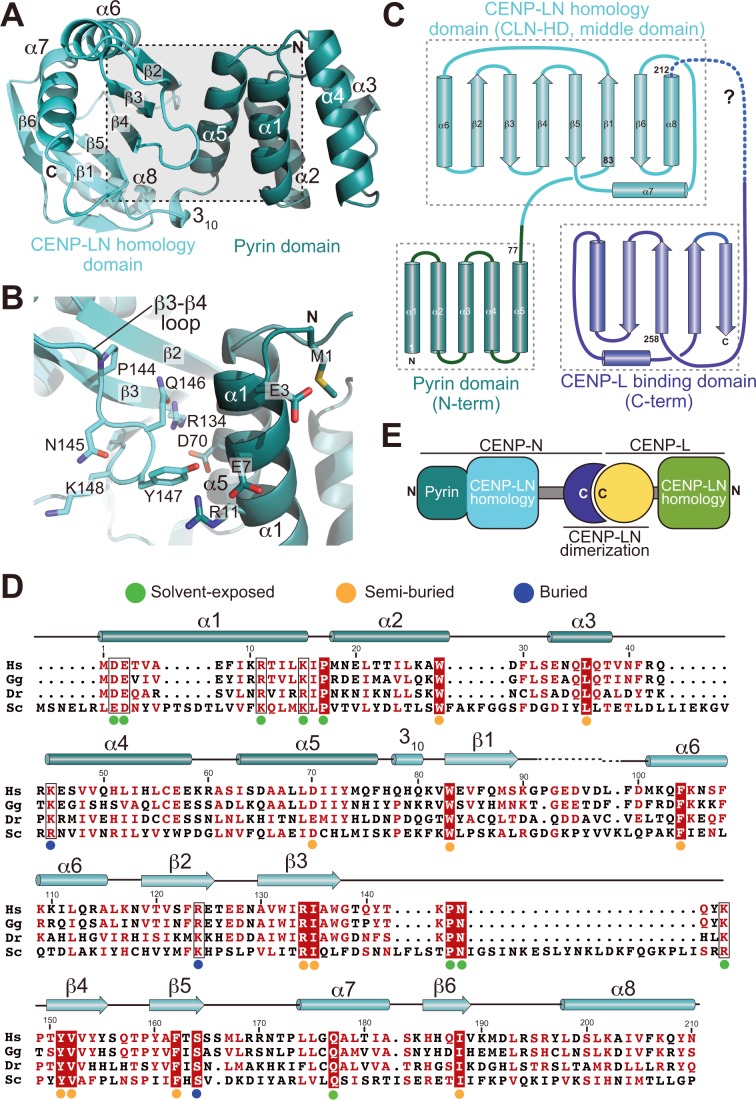
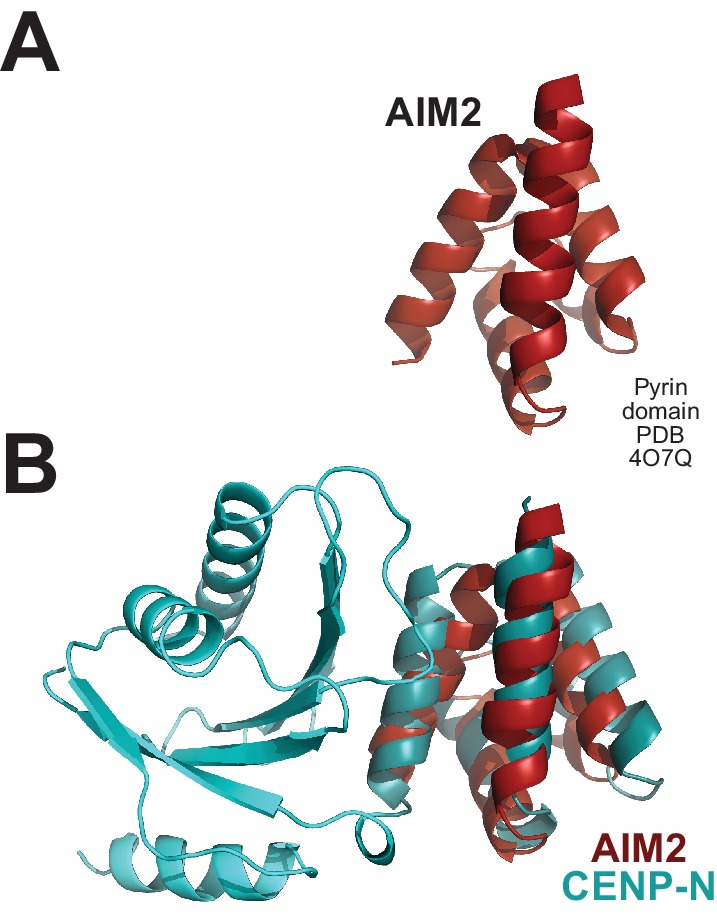
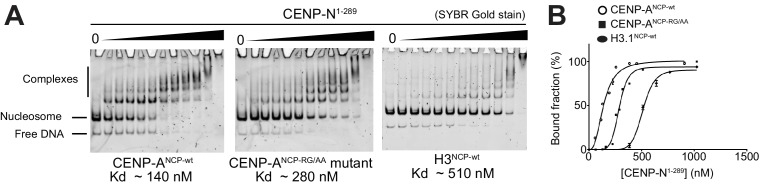
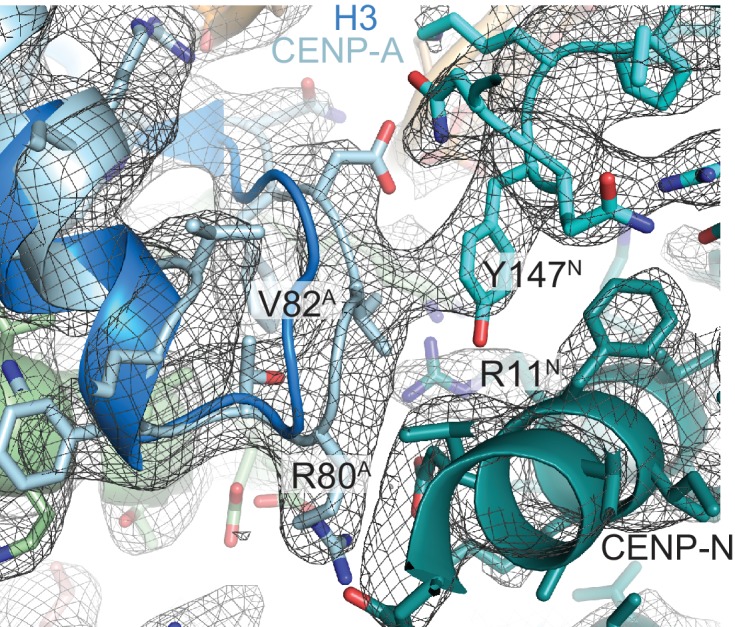
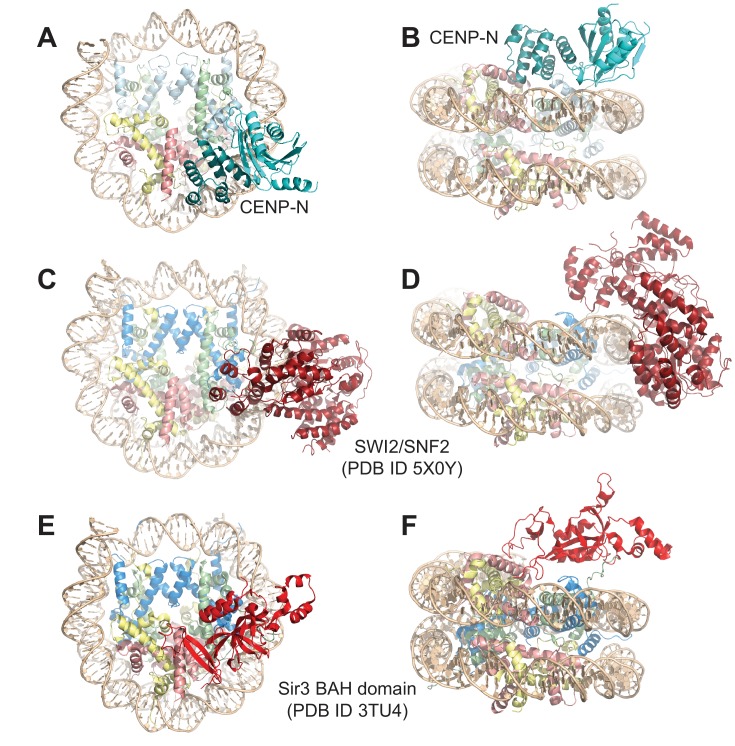
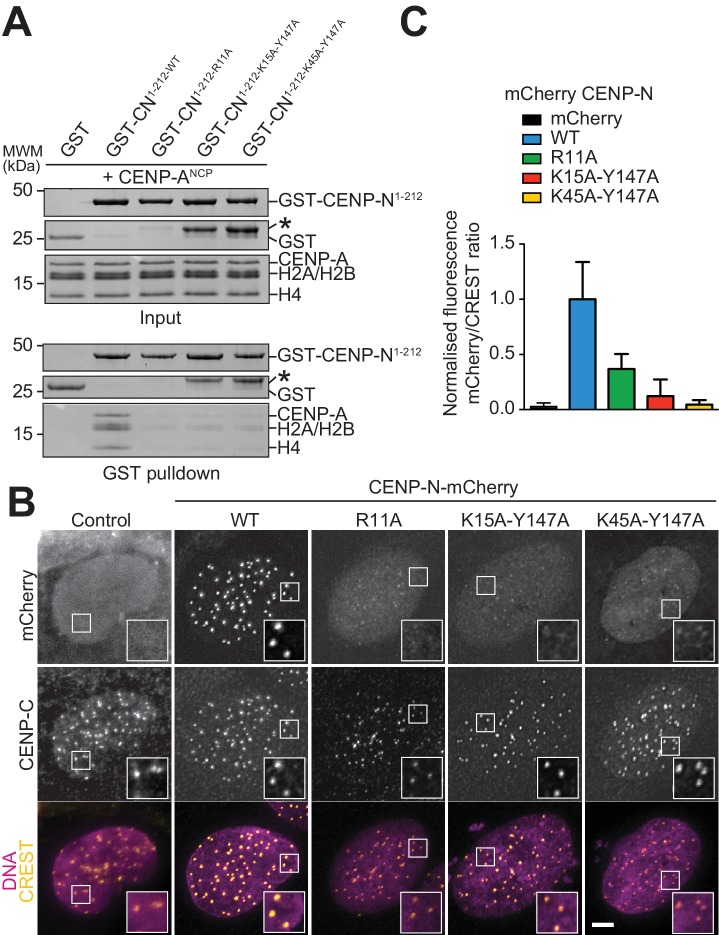
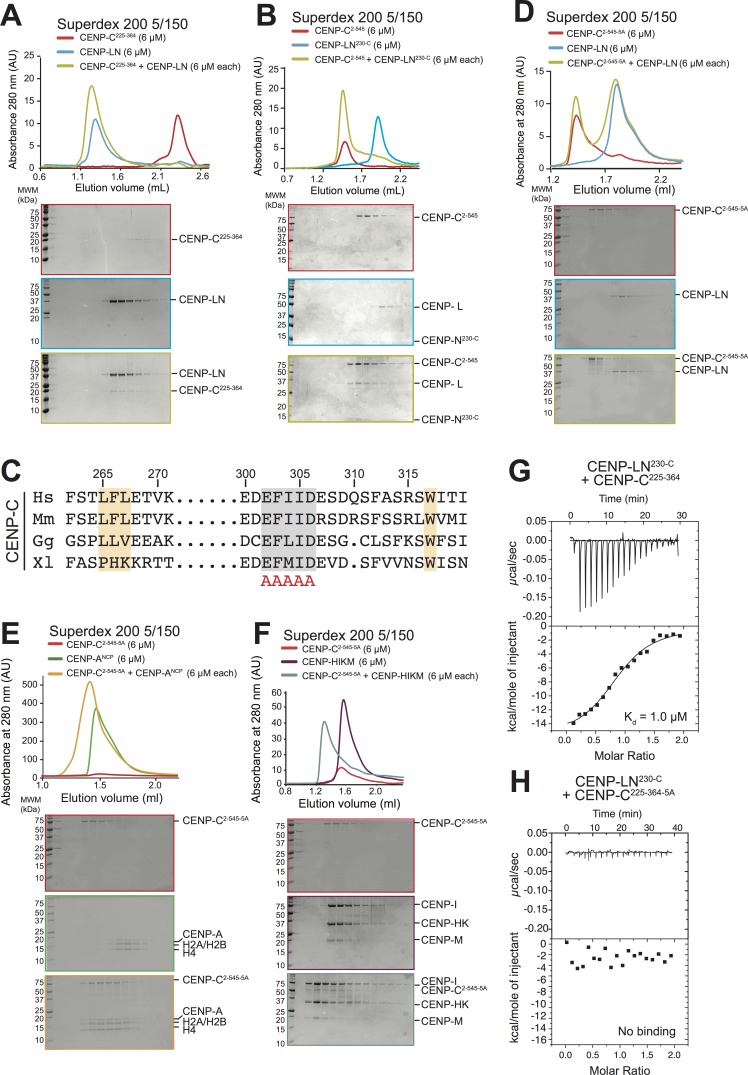
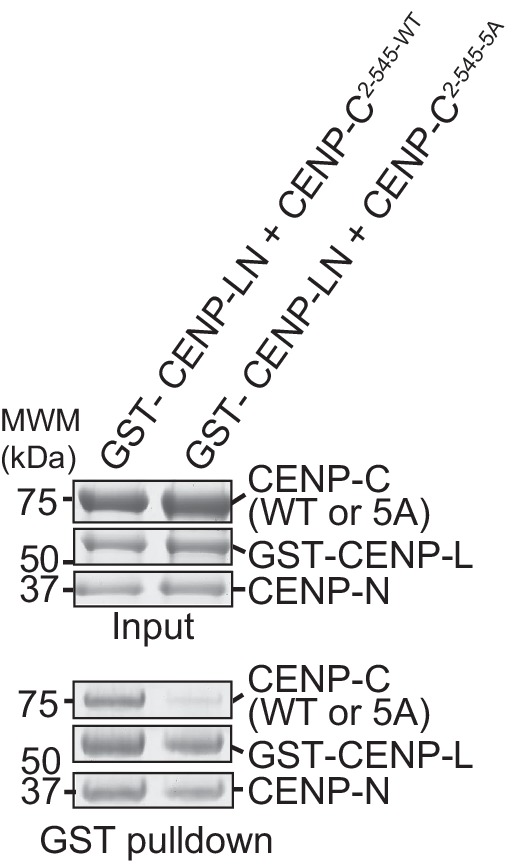
Centromere protein (CENP) A, a histone H3 variant, is a key epigenetic determinant of chromosome domains known as centromeres. Centromeres nucleate kinetochores, multi-subunit complexes that capture spindle microtubules to promote chromosome segregation during mitosis. Two kinetochore proteins, CENP-C and CENP-N, recognize CENP-A in the context of a rare CENP-A nucleosome. Here, we reveal the structural basis for the exquisite selectivity of CENP-N for centromeres. CENP-N uses charge and space complementarity to decode the L1 loop that is unique to CENP-A. It also engages in extensive interactions with a 15-base pair segment of the distorted nucleosomal DNA double helix, in a position predicted to exclude chromatin remodelling enzymes. Besides CENP-A, stable centromere recruitment of CENP-N requires a coincident interaction with a newly identified binding motif on nucleosome-bound CENP-C. Collectively, our studies clarify how CENP-N and CENP-C decode and stabilize the non-canonical CENP-A nucleosome to enforce epigenetic centromere specification and kinetochore assembly.
着丝粒蛋白 (CENP) A 是一种组蛋白 H3 变体,是一种关键的表观遗传决定因素,可将染色体结构域定义为着丝粒。着丝粒可启动动粒,这是一种多亚基复合物,可捕获纺锤体微管,以促进有丝分裂过程中的染色体分离。两种动粒蛋白 CENP-C 和 CENP-N 在罕见的 CENP-A 核小体背景下识别 CENP-A。在这里,我们揭示了 CENP-N 对着丝粒具有极高选择性的结构基础。CENP-N 利用电荷和空间互补性来解码 CENP-A 特有的 L1 环。它还与扭曲核小体 DNA 双螺旋的 15 个碱基对片段广泛相互作用,该位置预计可排除染色质重塑酶。除了 CENP-A,CENP-N 的稳定着丝粒募集还需要与核小体结合的 CENP-C 上新鉴定的结合基序同时发生相互作用。总的来说,我们的研究阐明了 CENP-N 和 CENP-C 如何解码和稳定非典型的 CENP-A 核小体,以强制建立表观遗传的着丝粒特征和动粒组装。