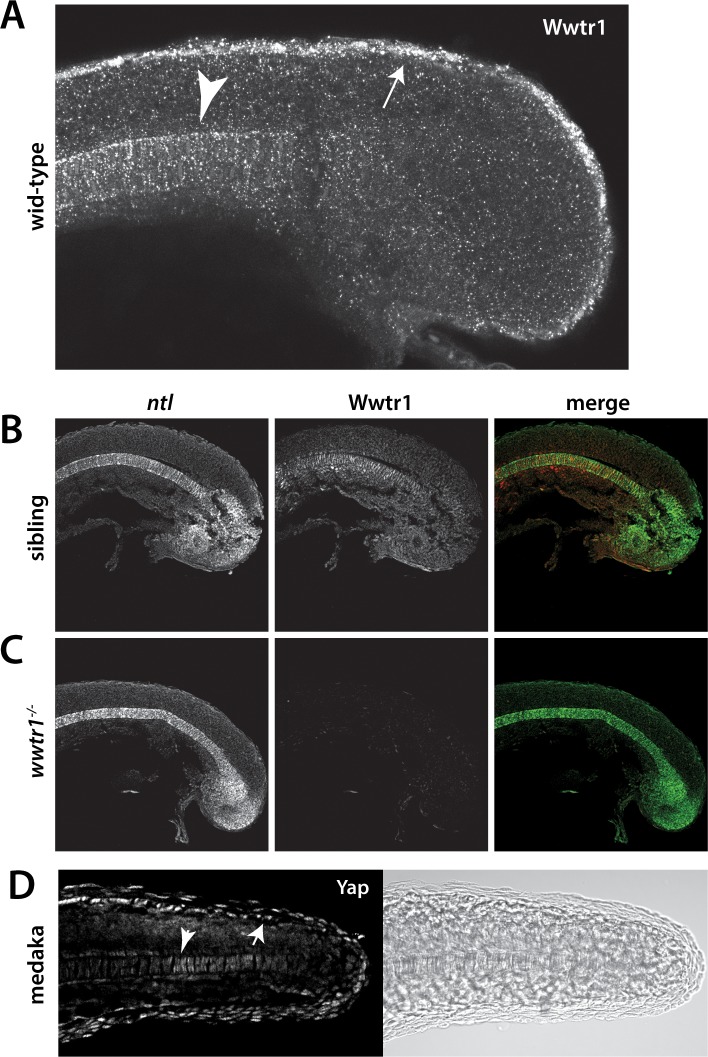
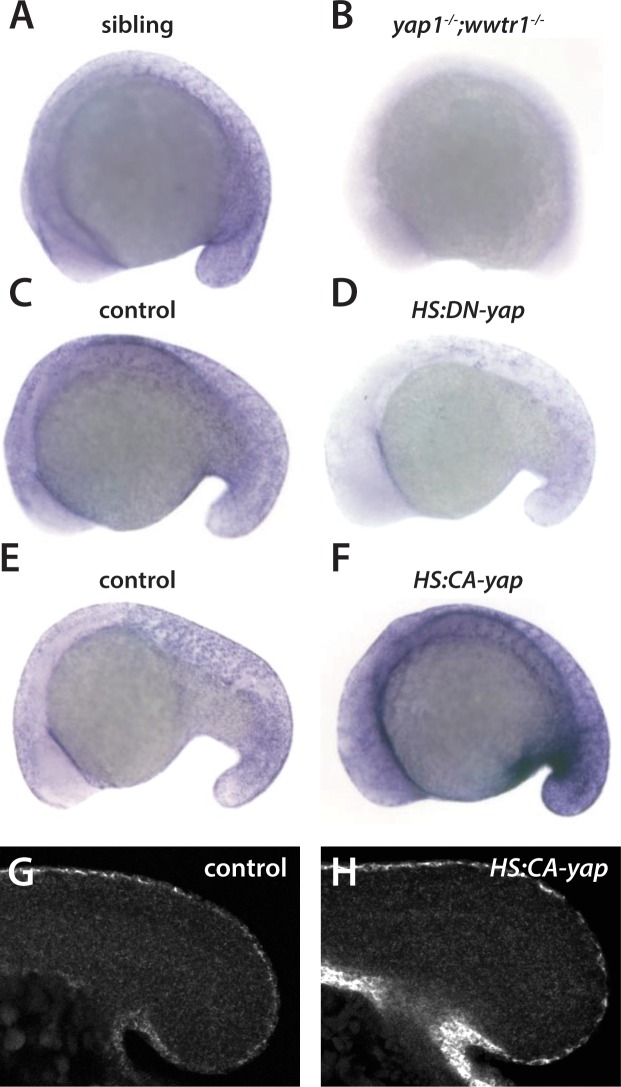
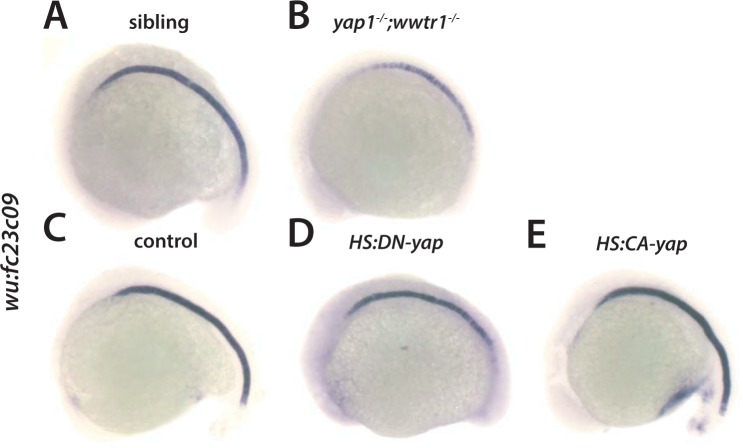
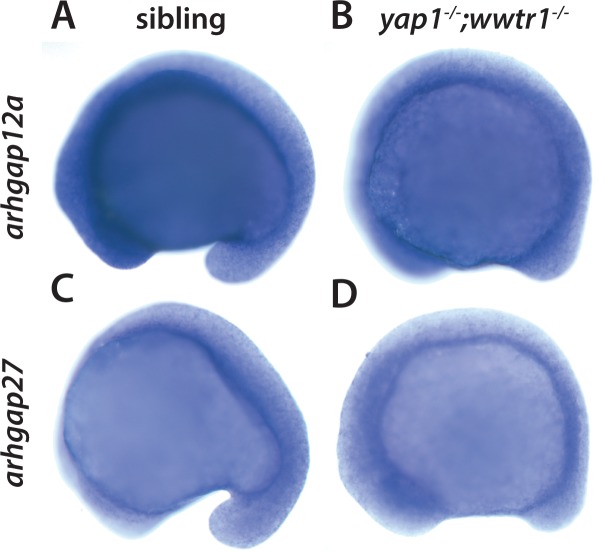
Department of Biochemistry, University of Washington, Seattle, United States.
Department of Developmental Genetics, Max Planck Institute for Heart and Lung Research, Bad Nauheim, Germany.
Elife. 2017 Dec 28;6:e31065. doi: 10.7554/eLife.31065.
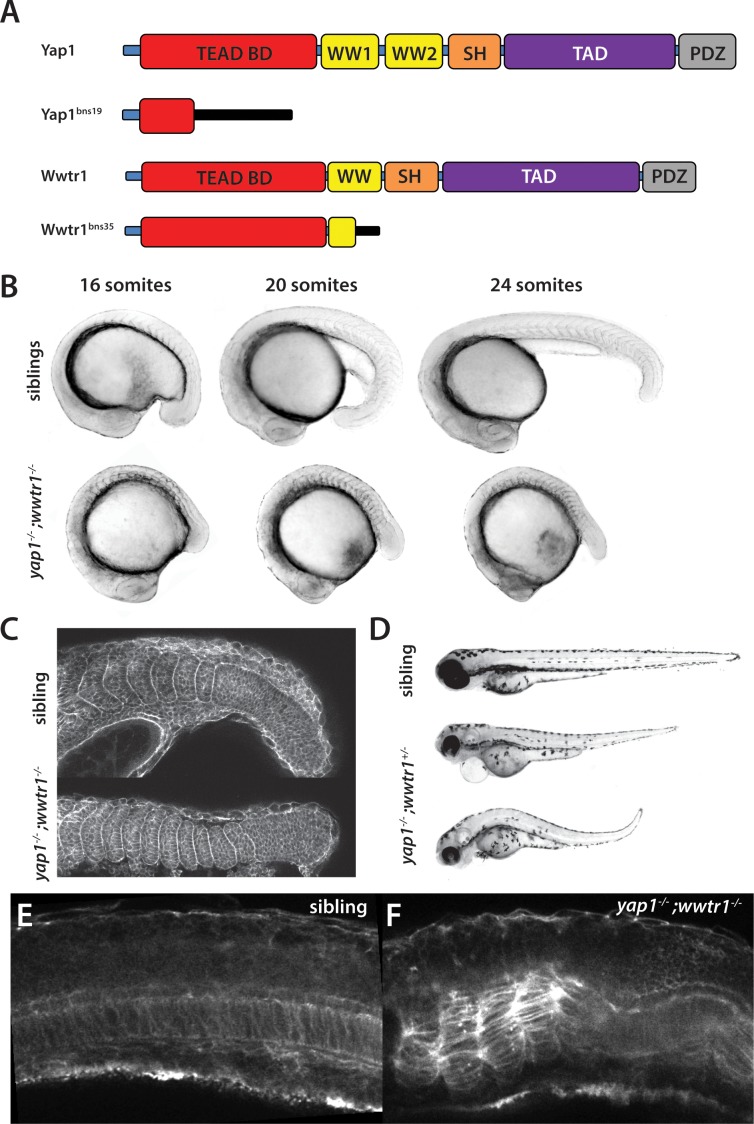
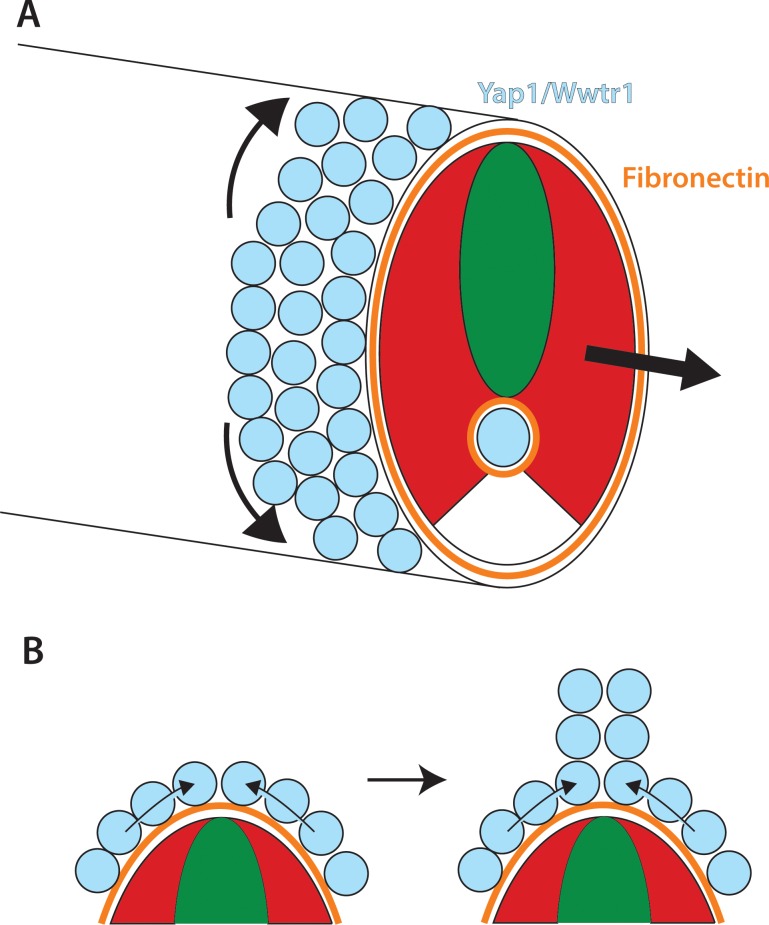
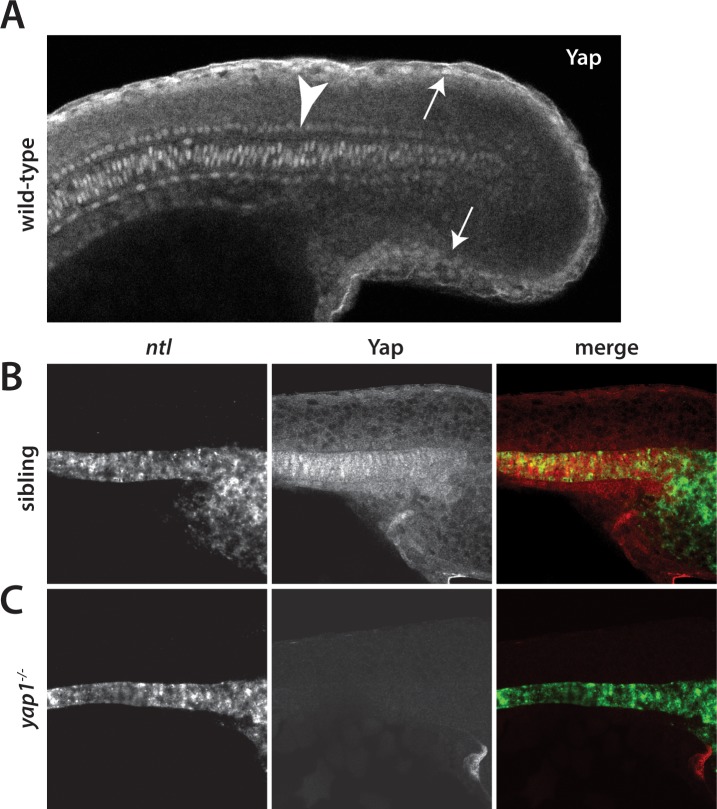
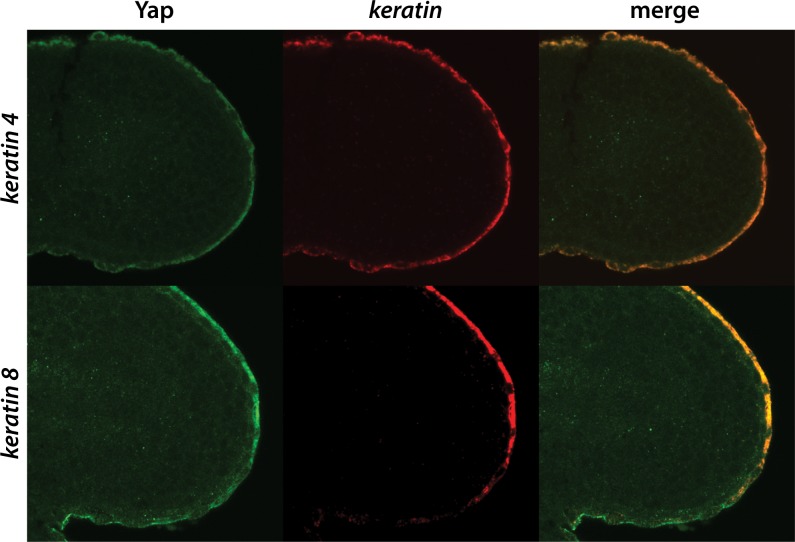
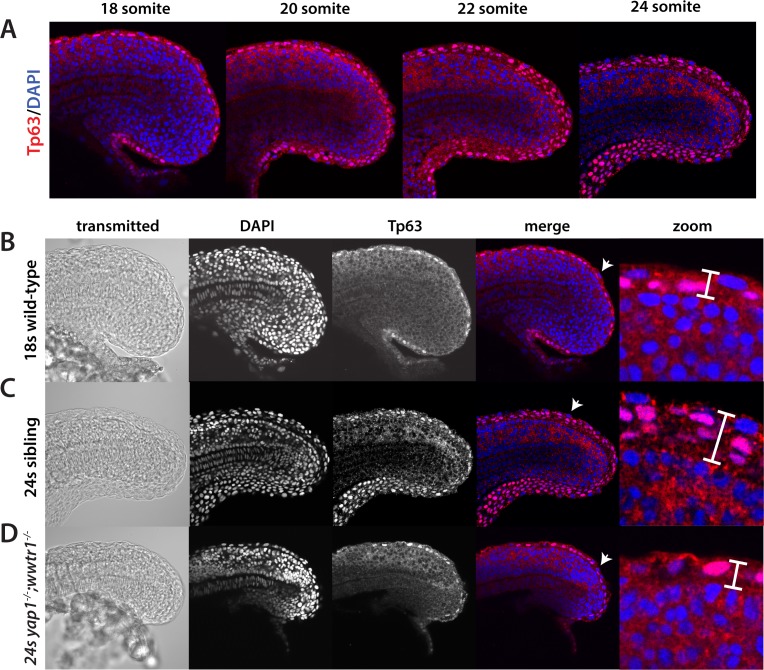
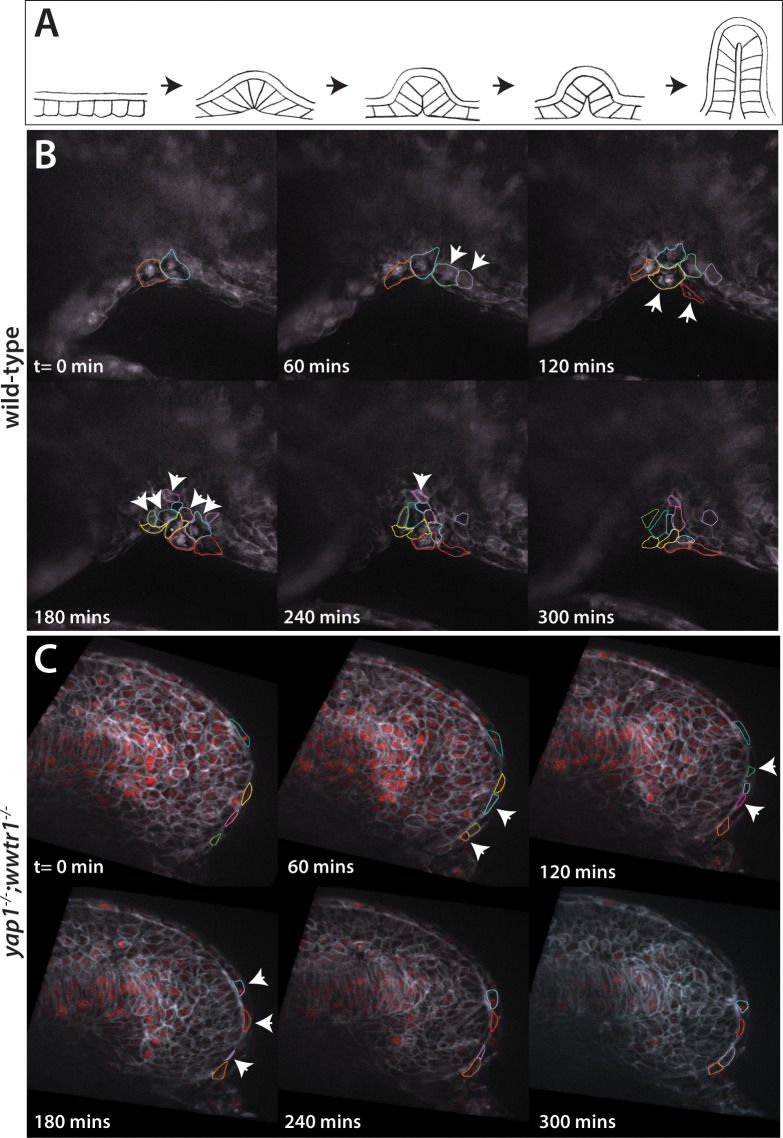
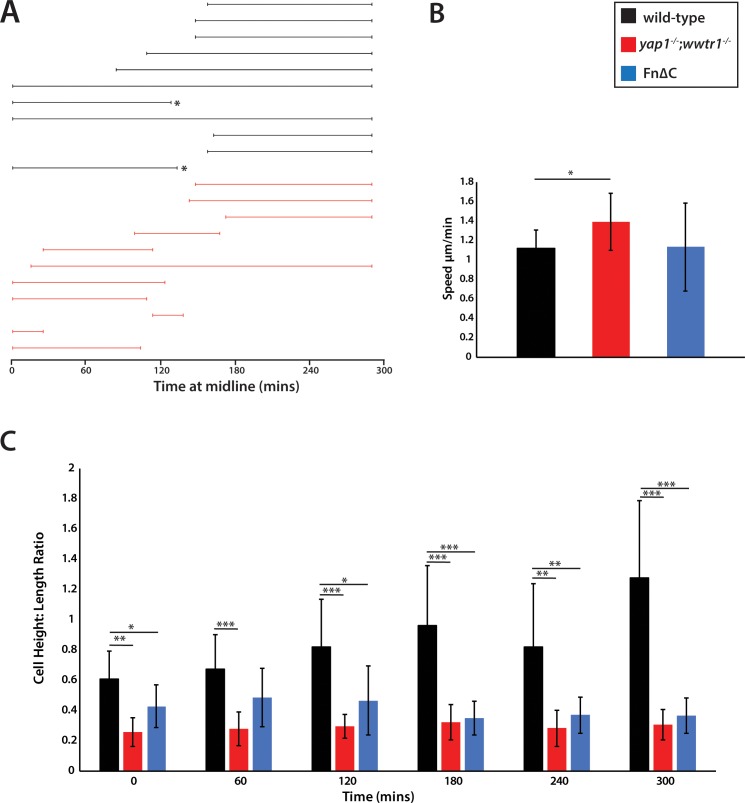
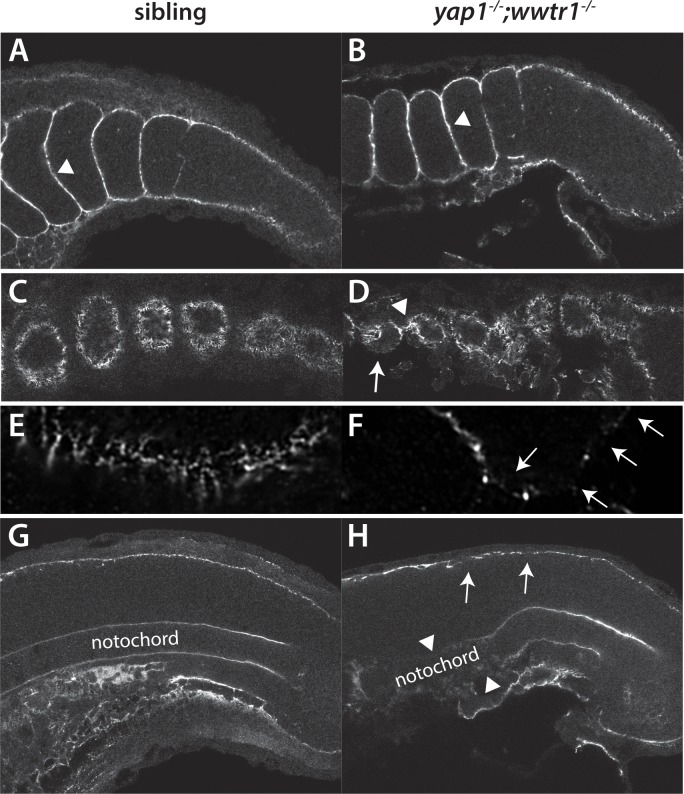
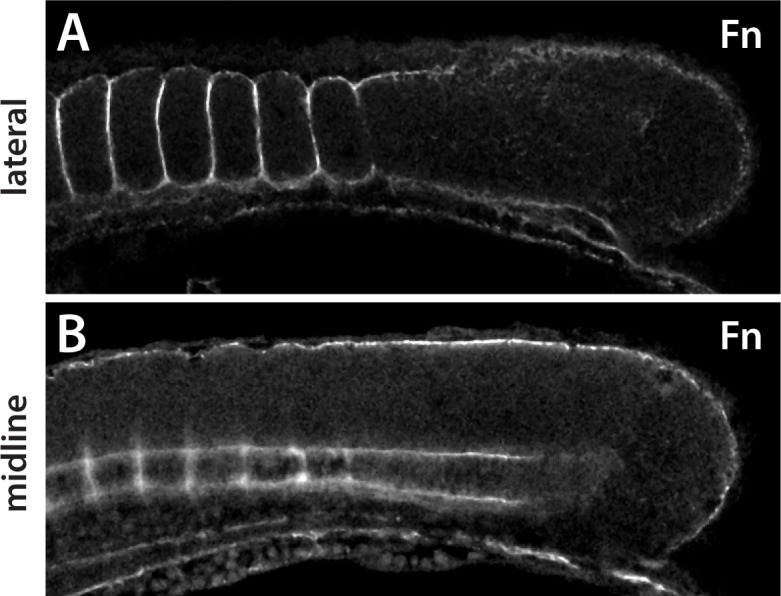
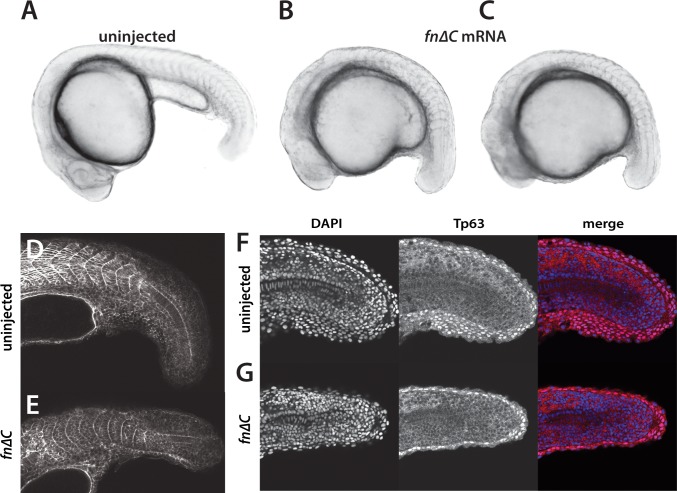
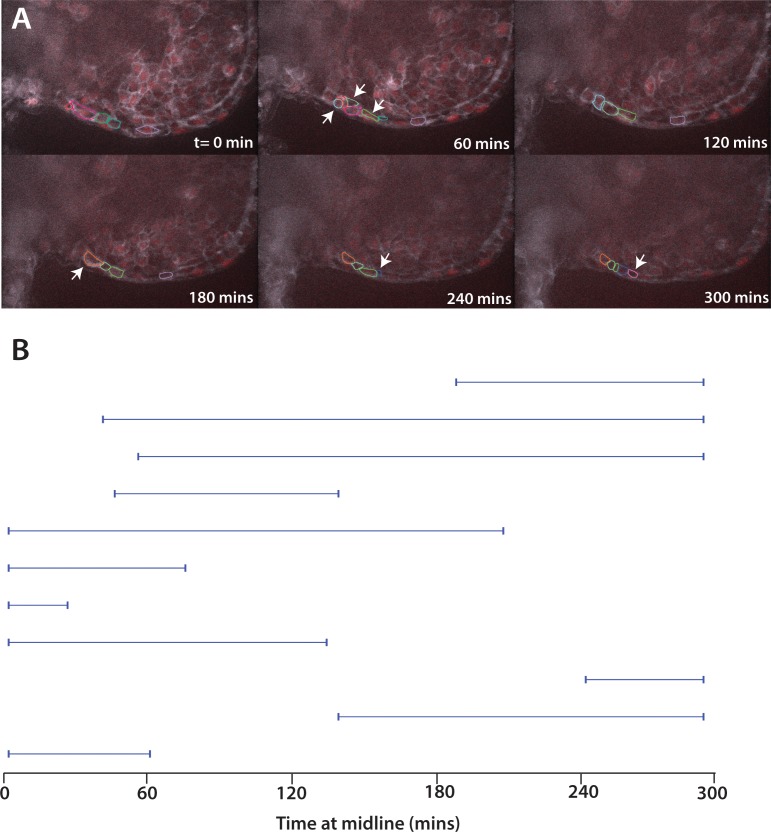
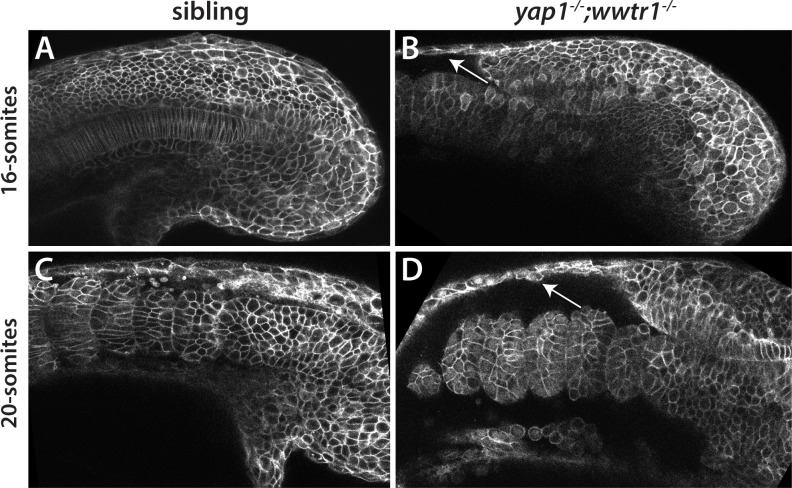
The vertebrate embryo undergoes a series of dramatic morphological changes as the body extends to form the complete anterior-posterior axis during the somite-forming stages. The molecular mechanisms regulating these complex processes are still largely unknown. We show that the Hippo pathway transcriptional coactivators Yap1 and Wwtr1 are specifically localized to the presumptive epidermis and notochord, and play a critical and unexpected role in posterior body extension by regulating Fibronectin assembly underneath the presumptive epidermis and surrounding the notochord. We further find that Yap1 and Wwtr1, also via Fibronectin, have an essential role in the epidermal morphogenesis necessary to form the initial dorsal and ventral fins, a process previously thought to involve bending of an epithelial sheet, but which we now show involves concerted active cell movement. Our results reveal how the Hippo pathway transcriptional program, localized to two specific tissues, acts to control essential morphological events in the vertebrate embryo.
脊椎动物胚胎在体节形成阶段,通过伸展身体形成完整的前-后轴,经历了一系列剧烈的形态变化。然而,调节这些复杂过程的分子机制在很大程度上仍不清楚。我们发现 Hippo 通路转录共激活因子 Yap1 和 Wwtr1 特异性定位于表皮和脊索的前体细胞,并通过调节表皮下和脊索周围的纤维连接蛋白组装,在胚胎后体延伸中发挥关键且意外的作用。我们进一步发现 Yap1 和 Wwtr1 也通过纤维连接蛋白在表皮形态发生中发挥重要作用,这是形成最初的背鳍和腹鳍所必需的过程,这个过程以前被认为涉及上皮片的弯曲,但我们现在表明,这涉及协调的主动细胞运动。我们的研究结果揭示了 Hippo 通路转录程序如何定位于两个特定组织,从而控制脊椎动物胚胎中的基本形态事件。