Padgett Leah R, Lentini Jenna M, Holmes Michael J, Stilger Krista L, Fu Dragony, Sullivan William J
Department of Pharmacology & Toxicology, Indiana University School of Medicine, Indianapolis, Indiana, United States of America.
Department of Biology, University of Rochester, Rochester, New York, United States of America.
PLoS One. 2018 Jan 2;13(1):e0189688. doi: 10.1371/journal.pone.0189688. eCollection 2018.
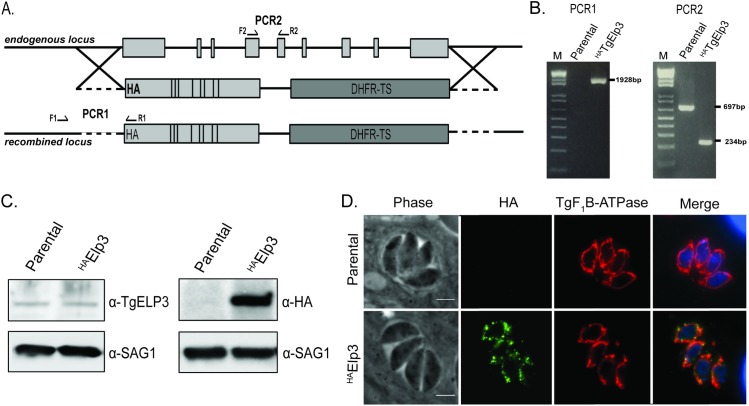
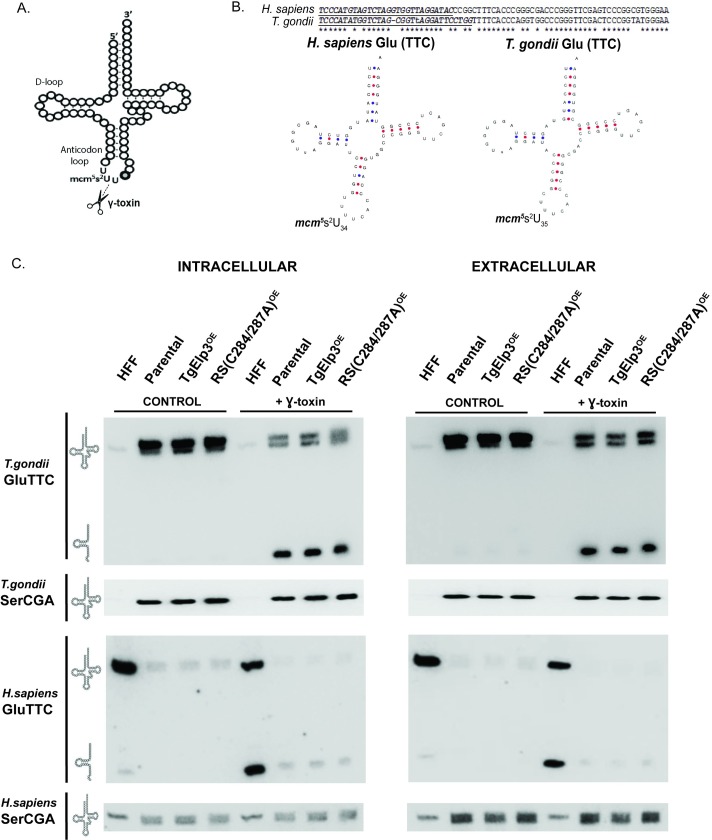
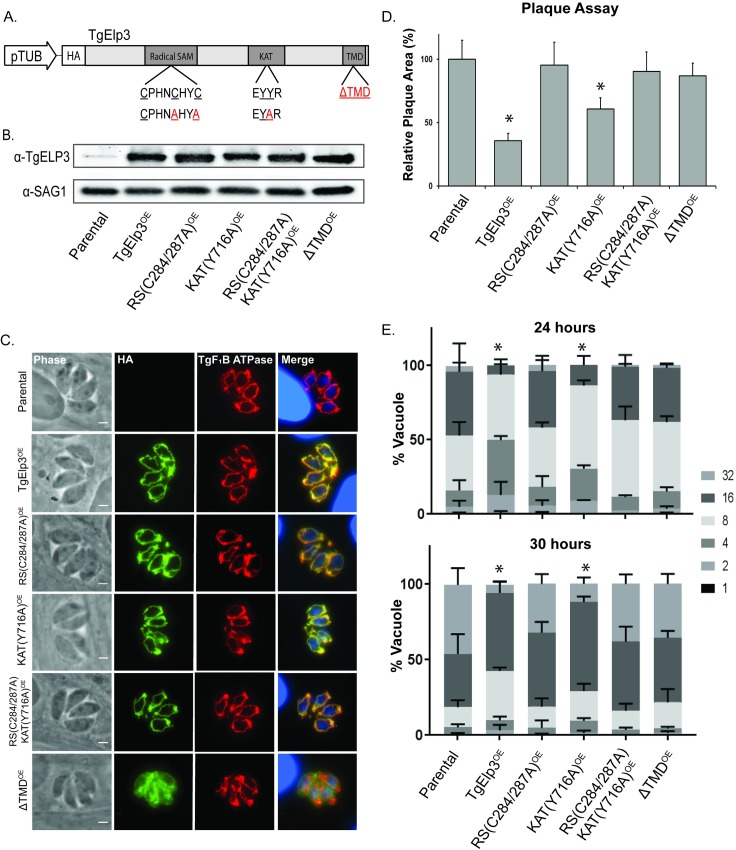
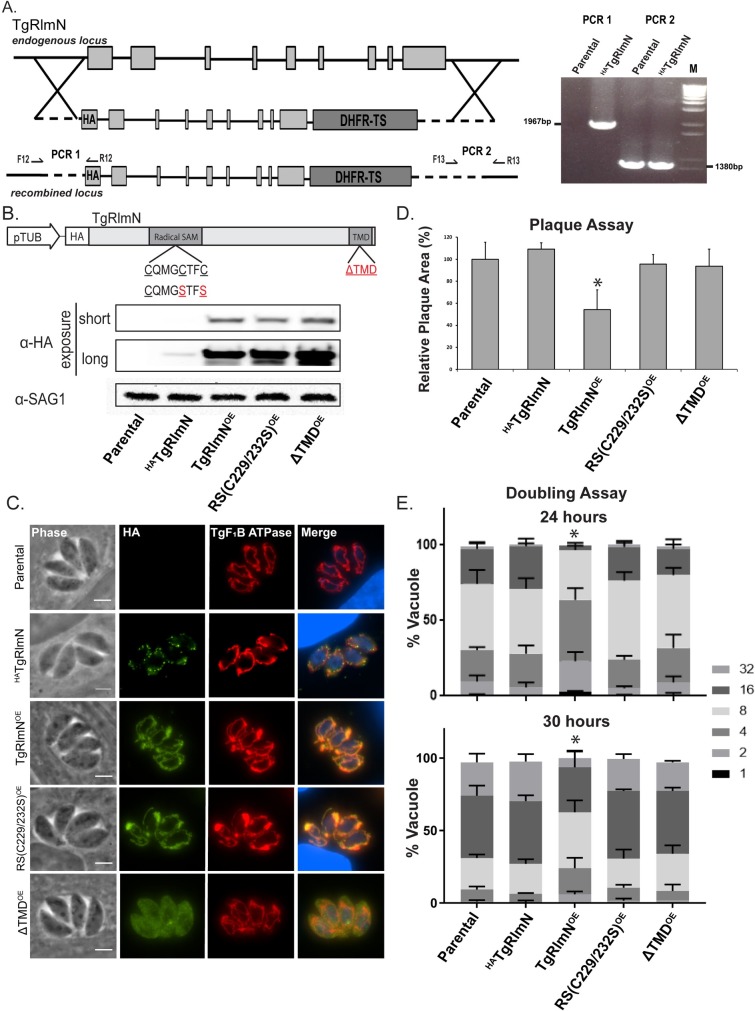
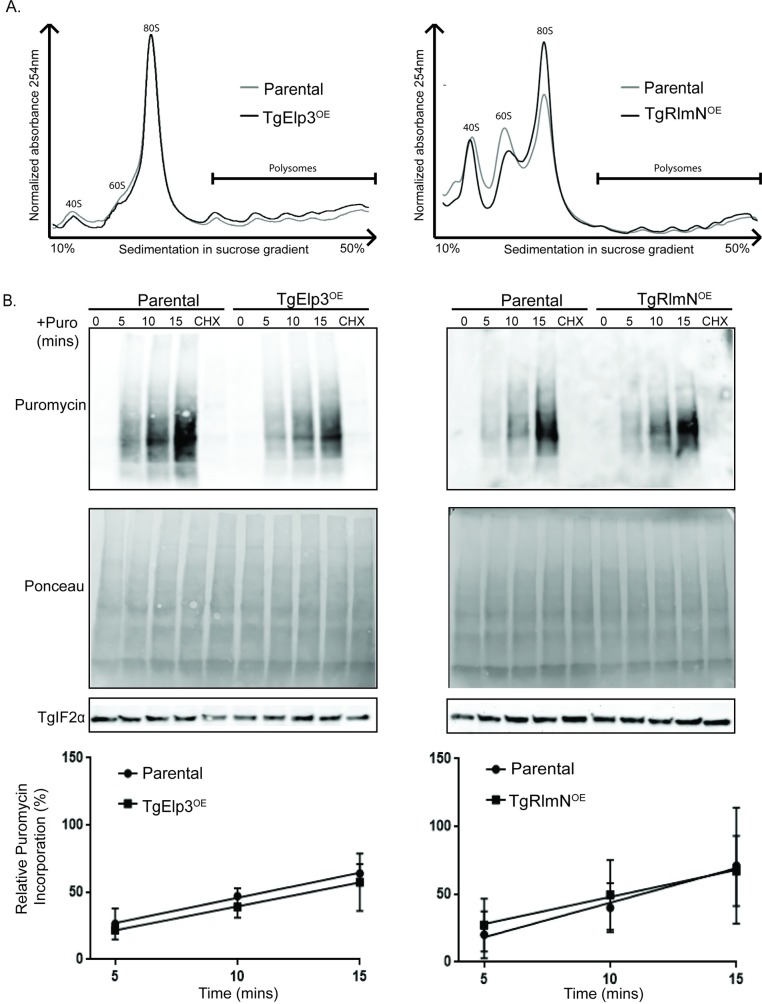
Radical S-adenosylmethionine (rSAM) enzymes use a 5'-deoxyadensyl 5'-radical to methylate a wide array of diverse substrates including proteins, lipids and nucleic acids. One such enzyme, Elongator protein-3 (TgElp3), is an essential protein in Toxoplasma gondii, a protozoan parasite that can cause life-threatening opportunistic disease. Unlike Elp3 homologues which are present in all domains of life, TgElp3 localizes to the outer mitochondrial membrane (OMM) via a tail-anchored trafficking mechanism in Toxoplasma. Intriguingly, we identified a second tail-anchored rSAM domain containing protein (TgRlmN) that also localizes to the OMM. The transmembrane domain (TMD) on Toxoplasma Elp3 and RlmN homologues is required for OMM localization and has not been seen beyond the chromalveolates. Both TgElp3 and TgRlmN contain the canonical rSAM amino acid sequence motif (CxxxCxxC) necessary to form the 4Fe-4S cluster required for tRNA modifications. In E. coli, RlmN is responsible for the 2-methlyadenosine (m2A) synthesis at purine 37 in tRNA while in S. cerevisiae, Elp3 is necessary for the formation of 5-methoxycarbonylmethyl-2-thiouridine (mcm5s2U) at the wobble tRNA position. To investigate why these two rSAM enzymes localize to the mitochondrion in Toxoplasma, and whether or not TgRlmN and TgElp3 possess tRNA methyltransferase activity, a series of mutational and biochemical studies were performed. Overexpression of either TgElp3 or TgRlmN resulted in a significant parasite replication defect, but overexpression was tolerated if either the TMD or rSAM domain was mutated. Furthermore, we show the first evidence that Toxoplasma tRNAGlu contains the mcm5s2U modification, which is the putative downstream product generated by TgElp3 activity.
自由基S-腺苷甲硫氨酸(rSAM)酶利用5'-脱氧腺苷5'-自由基对包括蛋白质、脂质和核酸在内的多种不同底物进行甲基化修饰。其中一种酶,延伸因子蛋白-3(TgElp3),是刚地弓形虫中的一种必需蛋白,刚地弓形虫是一种原生动物寄生虫,可引发危及生命的机会性疾病。与存在于所有生命域中的Elp3同源物不同,TgElp3通过刚地弓形虫中一种尾锚定转运机制定位于线粒体外膜(OMM)。有趣的是,我们鉴定出另一种含有尾锚定rSAM结构域的蛋白(TgRlmN),它也定位于OMM。刚地弓形虫Elp3和RlmN同源物上的跨膜结构域(TMD)是OMM定位所必需的,并且在色素体生物之外尚未发现。TgElp3和TgRlmN都包含形成tRNA修饰所需的4Fe-4S簇所必需的典型rSAM氨基酸序列基序(CxxxCxxC)。在大肠杆菌中,RlmN负责tRNA中嘌呤37位的2-甲基腺苷(m2A)合成,而在酿酒酵母中,Elp3是tRNA摆动位置处5-甲氧基羰基甲基-2-硫代尿苷(mcm5s2U)形成所必需的。为了研究这两种rSAM酶为何在刚地弓形虫中定位于线粒体,以及TgRlmN和TgElp3是否具有tRNA甲基转移酶活性,我们进行了一系列突变和生化研究。TgElp3或TgRlmN的过表达导致明显的寄生虫复制缺陷,但如果TMD或rSAM结构域发生突变,则过表达可被耐受。此外,我们首次证明刚地弓形虫tRNAGlu含有mcm5s2U修饰,这是TgElp3活性产生的假定下游产物。