Khalil Shadi, Holy Maja, Grado Stephen, Fleming Robert, Kurita Ryo, Nakamura Yukio, Goldfarb Adam
Department of Pathology, University of Virginia School of Medicine, Charlottesville, VA.
Department of Pediatrics, Saint Louis University School of Medicine, St. Louis, MO.
Blood Adv. 2017 Jun 27;1(15):1181-1194. doi: 10.1182/bloodadvances.2016003772.
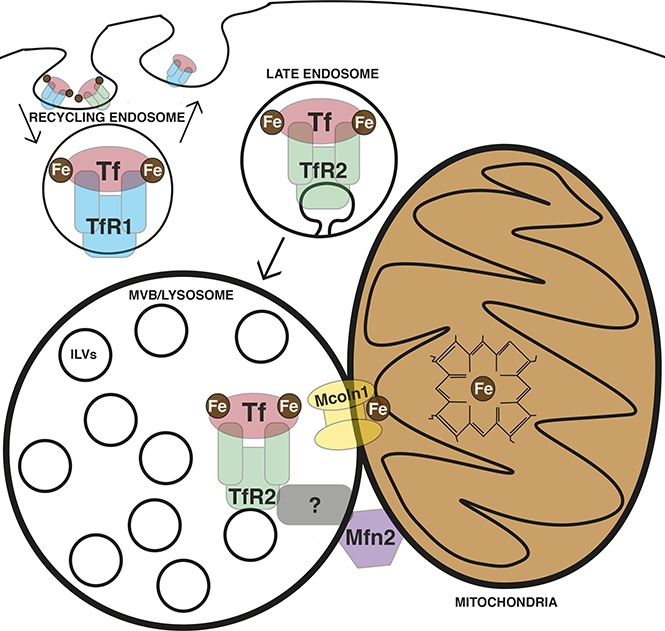
Erythroid progenitors are the largest consumers of iron in the human body. In these cells, a high flux of iron must reach the mitochondrial matrix to form sufficient heme to support hemoglobinization. Canonical erythroid iron trafficking occurs via the first transferrin receptor (TfR1)-mediated endocytosis of diferric-transferrin into recycling endosomes, where ferric iron is released, reduced, and exported to the cytosol via DMT1. However, mice lacking TfR1 or DMT1 demonstrate residual erythropoiesis, suggesting additional pathways for iron use. How iron moves from endosomes to mitochondria is incompletely understood, with both cytosolic chaperoning and "kiss and run" interorganelle transfer implicated. TfR2, in contrast to its paralog TfR1, has established roles in iron sensing, but not iron uptake. Recently, mice with marrow-selective TfR2 deficiency were found to exhibit microcytosis, suggesting TfR2 may also contribute to erythroid hemoglobinization. In this study, we identify alternative trafficking, in which TfR2 mediates lysosomal transferrin delivery. Imaging studies reveal an erythroid lineage-specific organelle arrangement consisting of a focal lysosomal cluster surrounded by a nest of mitochondria, with direct contacts between these 2 organelles. Erythroid TfR2 deficiency yields aberrant mitochondrial morphology, implicating TfR2-dependent transferrin trafficking in mitochondrial maintenance. Human shares a lineage- and stage-specific expression pattern with , encoding a lysosomal iron channel, and , encoding a protein mediating organelle contacts. Functional studies reveal these latter factors to be involved in mitochondrial regulation and erythroid differentiation, with Mfn2 required for mitochondrial-lysosomal contacts. These findings identify a new pathway for erythroid iron trafficking involving TfR2-mediated lysosomal delivery followed by interorganelle transfer to mitochondria.
红系祖细胞是人体中最大的铁消耗者。在这些细胞中,大量的铁必须进入线粒体基质以形成足够的血红素,从而支持血红蛋白化。经典的红系铁转运过程是通过第一个转铁蛋白受体(TfR1)介导的双铁转铁蛋白内吞进入循环内体,在那里三价铁被释放、还原,并通过二价金属离子转运体1(DMT1)输出到细胞质中。然而,缺乏TfR1或DMT1的小鼠仍表现出残余的红细胞生成,这表明存在其他铁利用途径。铁如何从内体转移到线粒体尚不完全清楚,胞质伴侣蛋白和“亲吻与跑”的细胞器间转移都被认为与之有关。与它的同源物TfR1不同,TfR2在铁感应而非铁摄取方面发挥作用。最近发现,骨髓选择性TfR2缺陷的小鼠表现出小红细胞症,这表明TfR2也可能有助于红系血红蛋白化。在本研究中,我们确定了另一种转运方式,即TfR2介导溶酶体转铁蛋白递送。成像研究揭示了一种红系谱系特异性的细胞器排列,由一个局灶性溶酶体簇被一群线粒体包围组成,这两种细胞器之间存在直接接触。红系TfR2缺陷导致线粒体形态异常,这表明TfR2依赖的转铁蛋白转运在线粒体维持中起作用。人类与编码溶酶体铁通道的 和编码介导细胞器接触的蛋白质的 具有谱系和阶段特异性的表达模式。功能研究表明,后两个因子参与线粒体调节和红系分化,线粒体-溶酶体接触需要线粒体融合蛋白2(Mfn2)。这些发现确定了一种新的红系铁转运途径,涉及TfR2介导的溶酶体递送,随后进行细胞器间转移至线粒体。