Rudolf Virchow Center for Experimental Biomedicine, University of Würzburg, 97080 Würzburg, Germany.
Helen L. and Martin S. Kimmel Center for Biology and Medicine, Skirball Institute of Biomolecular Medicine, New York University School of Medicine, New York, NY 10016.
Proc Natl Acad Sci U S A. 2018 Feb 6;115(6):E1127-E1136. doi: 10.1073/pnas.1714085115. Epub 2018 Jan 24.
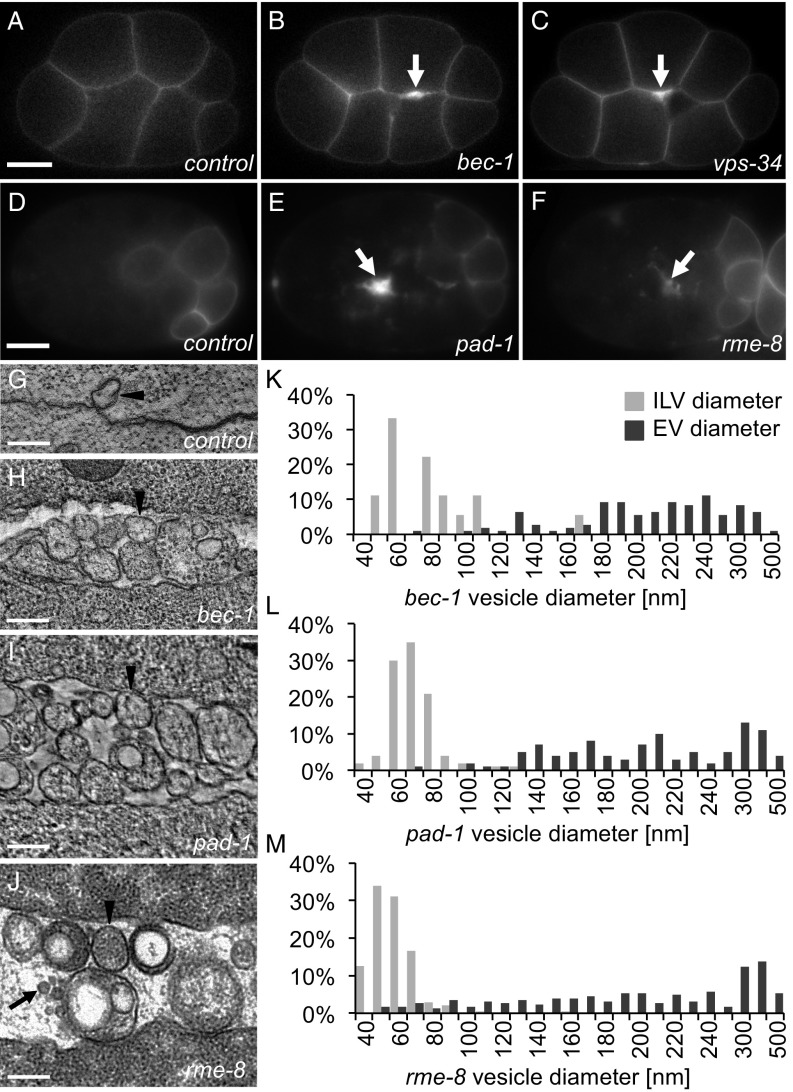
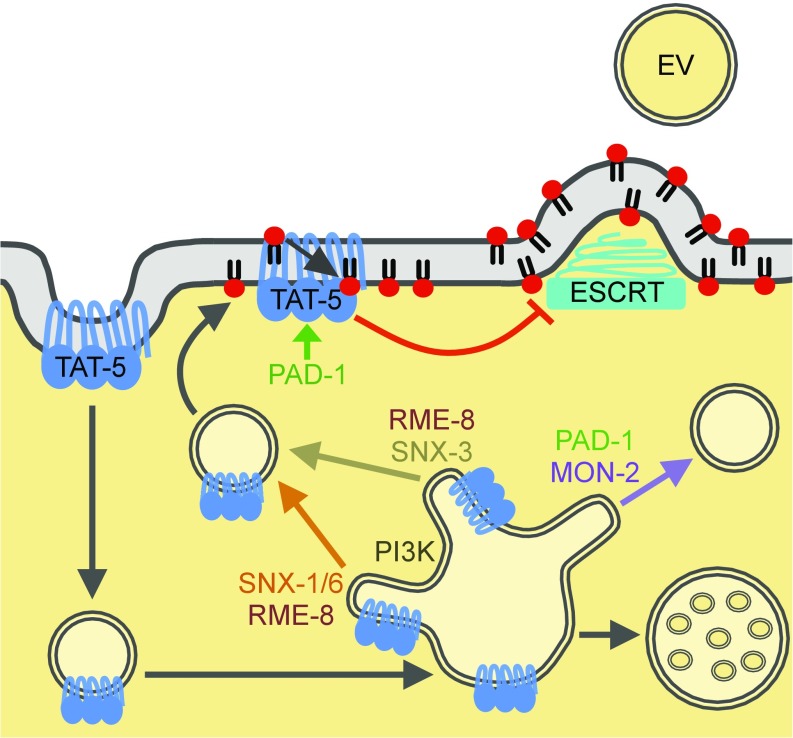
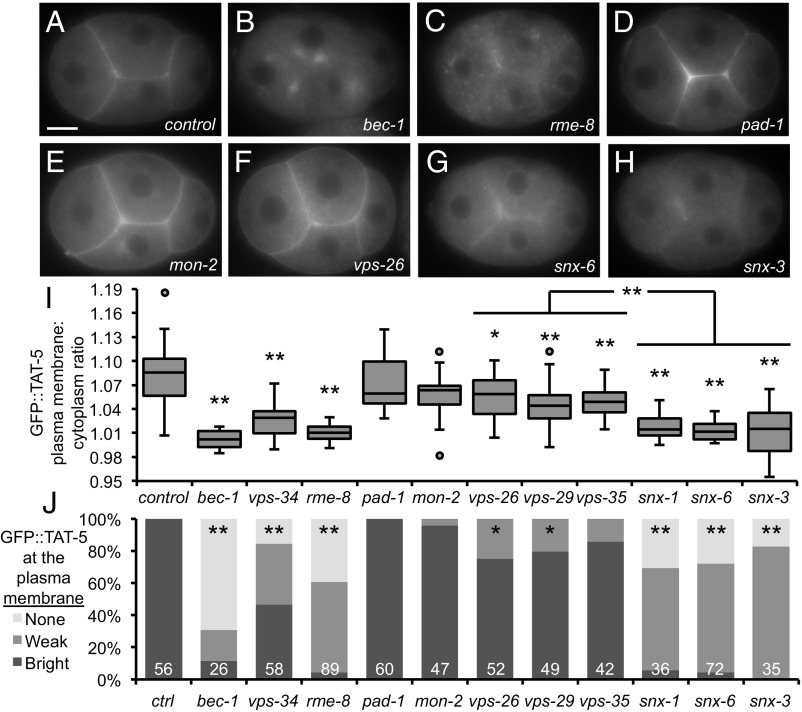
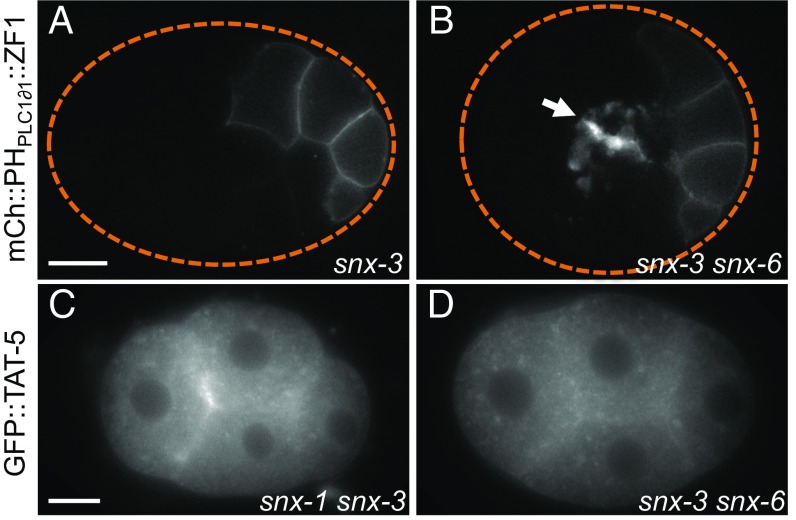
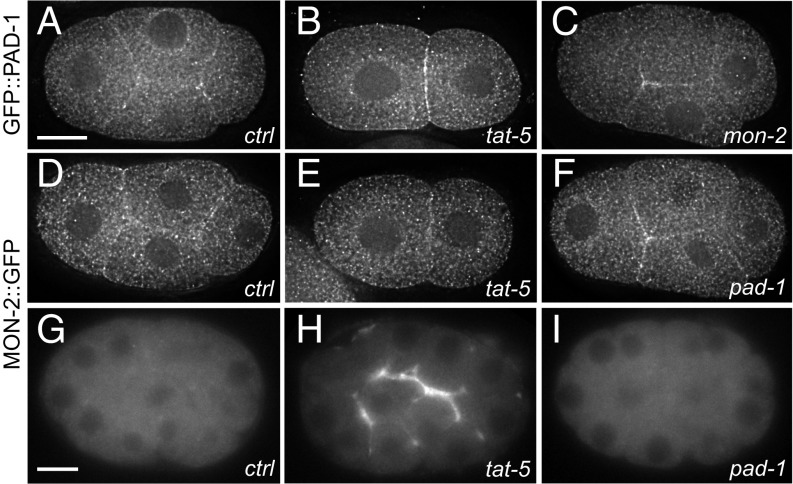
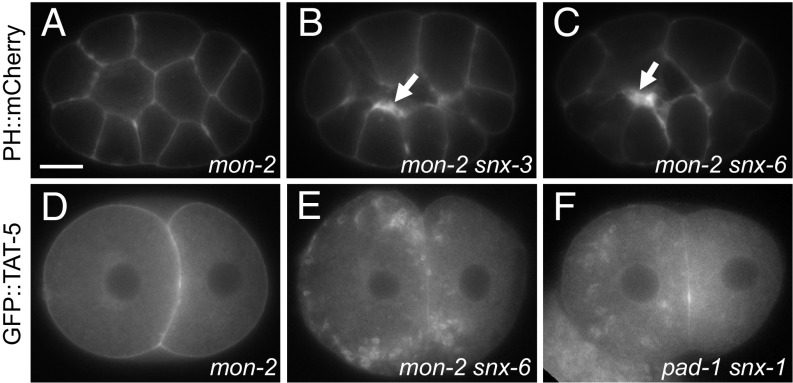
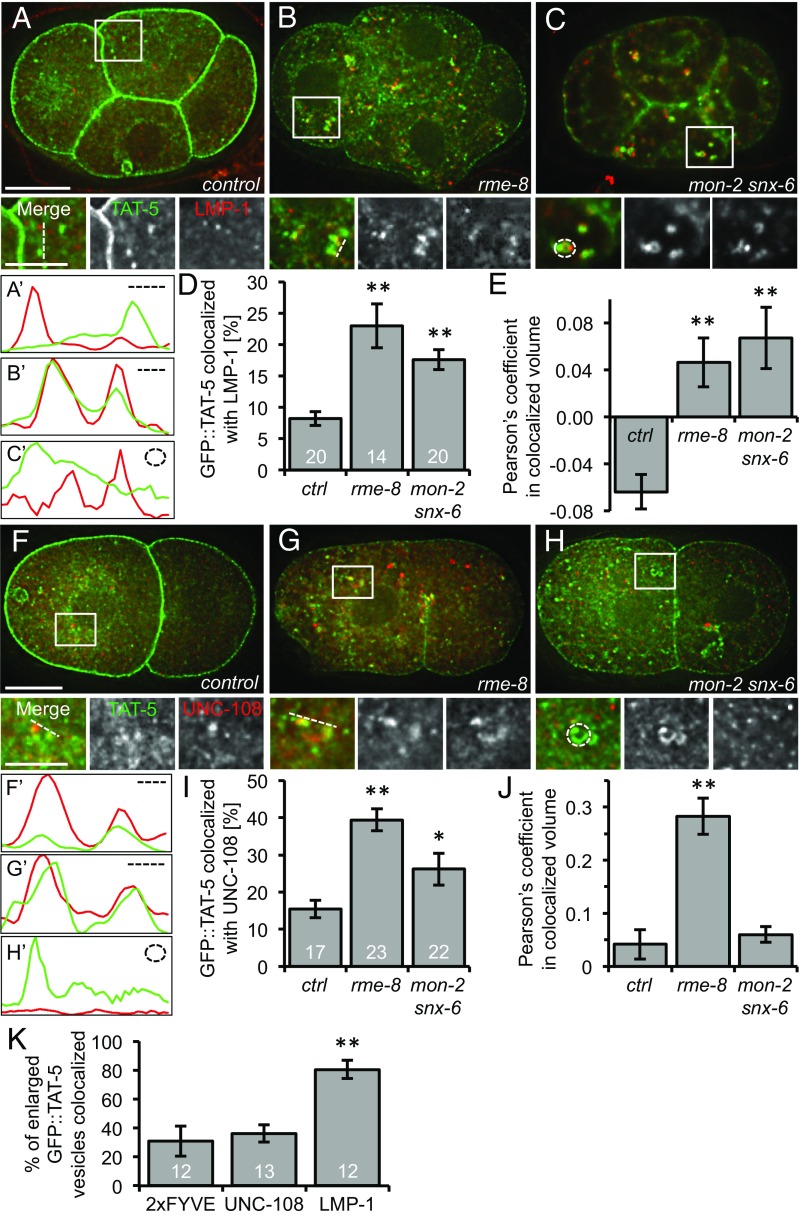
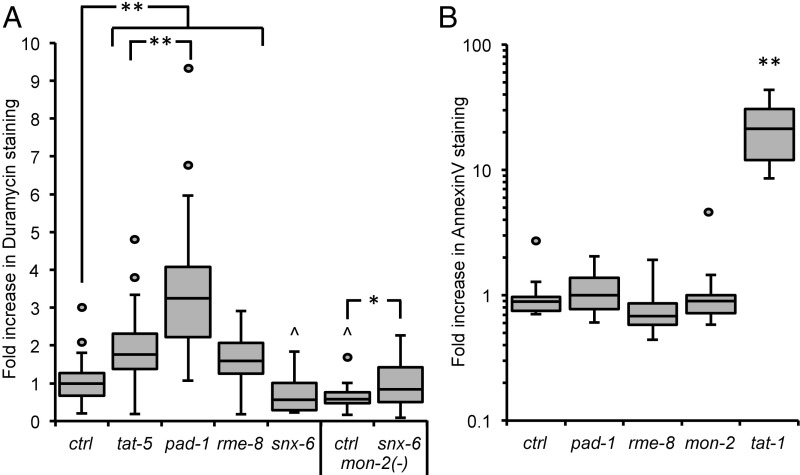
Cells release extracellular vesicles (EVs) that mediate intercellular communication and repair damaged membranes. Despite the pleiotropic functions of EVs in vitro, their in vivo function is debated, largely because it is unclear how to induce or inhibit their formation. In particular, the mechanisms of EV release by plasma membrane budding or ectocytosis are poorly understood. We previously showed that TAT-5 phospholipid flippase activity maintains the asymmetric localization of the lipid phosphatidylethanolamine (PE) in the plasma membrane and inhibits EV budding by ectocytosis in However, no proteins that inhibit ectocytosis upstream of TAT-5 were known. Here, we identify TAT-5 regulators associated with retrograde endosomal recycling: PI3Kinase VPS-34, Beclin1 homolog BEC-1, DnaJ protein RME-8, and the uncharacterized Dopey homolog PAD-1. PI3Kinase, RME-8, and semiredundant sorting nexins are required for the plasma membrane localization of TAT-5, which is important to maintain PE asymmetry and inhibit EV release. PAD-1 does not directly regulate TAT-5 localization, but is required for the lipid flipping activity of TAT-5. PAD-1 also has roles in endosomal trafficking with the GEF-like protein MON-2, which regulates PE asymmetry and EV release redundantly with sorting nexins independent of the core retromer. Thus, in addition to uncovering redundant intracellular trafficking pathways, our study identifies additional proteins that regulate EV release. This work pinpoints TAT-5 and PE as key regulators of plasma membrane budding, further supporting the model that PE externalization drives ectocytosis.
细胞释放细胞外囊泡 (EVs),介导细胞间通讯和修复受损的膜。尽管 EVs 在体外具有多种功能,但它们在体内的功能仍存在争议,主要是因为尚不清楚如何诱导或抑制其形成。特别是,质膜出芽或胞吐作用释放 EV 的机制还了解甚少。我们之前表明,TAT-5 磷脂翻转酶活性维持了质膜中脂质磷脂酰乙醇胺 (PE) 的不对称定位,并通过胞吐作用抑制了 EV 的出芽,然而,在 TAT-5 之前没有已知的抑制胞吐作用的蛋白质。在这里,我们确定了与逆行内体回收相关的 TAT-5 调节剂:PI3 激酶 VPS-34、Beclin1 同源物 BEC-1、DnaJ 蛋白 RME-8 和未表征的 Dopey 同源物 PAD-1。PI3 激酶、RME-8 和半冗余分选连接蛋白对于 TAT-5 在质膜上的定位是必需的,这对于维持 PE 不对称性和抑制 EV 释放很重要。PAD-1 不直接调节 TAT-5 的定位,但对于 TAT-5 的脂质翻转活性是必需的。PAD-1 还与 GEF 样蛋白 MON-2 一起在内体运输中发挥作用,它与分选连接蛋白一起独立于核心逆行体调节 PE 不对称性和 EV 释放,这与核心逆行体无关。因此,除了揭示冗余的细胞内运输途径外,我们的研究还确定了其他调节 EV 释放的蛋白质。这项工作指出 TAT-5 和 PE 是质膜出芽的关键调节剂,进一步支持了 PE 外化驱动胞吐作用的模型。