Research Center for Radiation Emergency Medicine, National Institute of Radiological Sciences (NIRS), Anagawa, Inage-ku, Chiba, Japan.
Program in Chemistry and Life Science, Department of Interdisciplinary Science and Engineering, School of Science and Engineering, Meisei University, Hodokubo, Hino-shi, Tokyo, Japan.
PLoS Genet. 2018 Mar 28;14(3):e1007277. doi: 10.1371/journal.pgen.1007277. eCollection 2018 Mar.
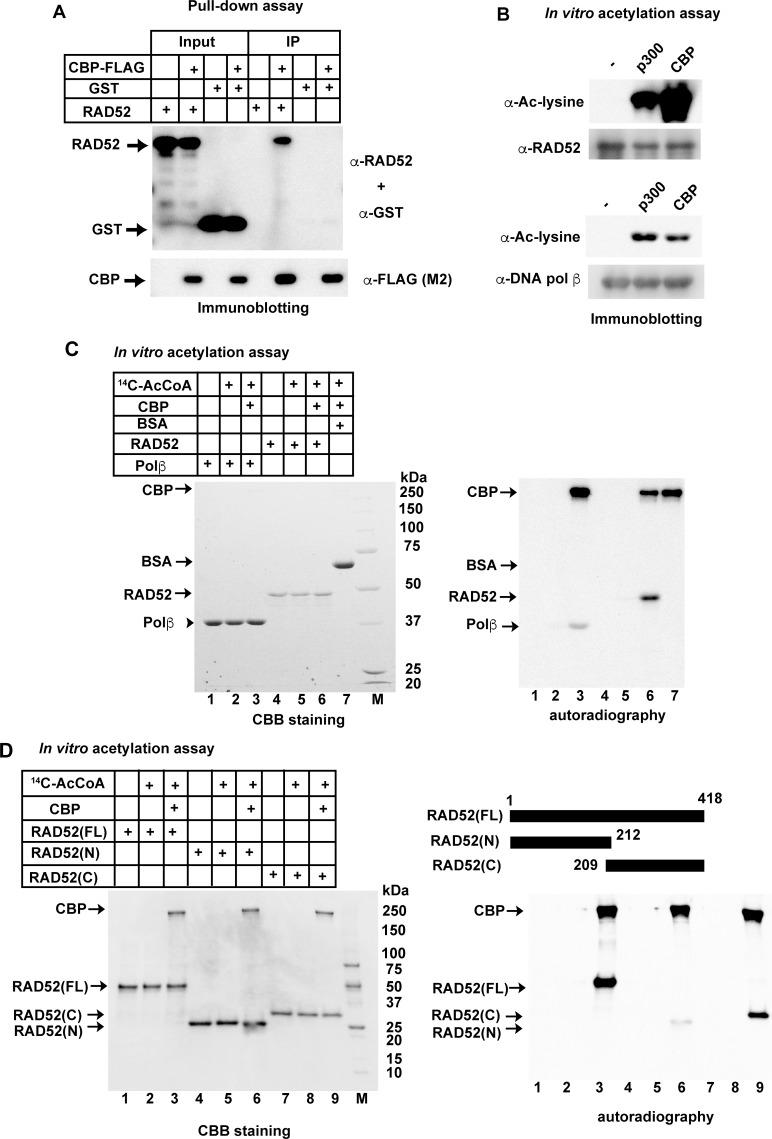
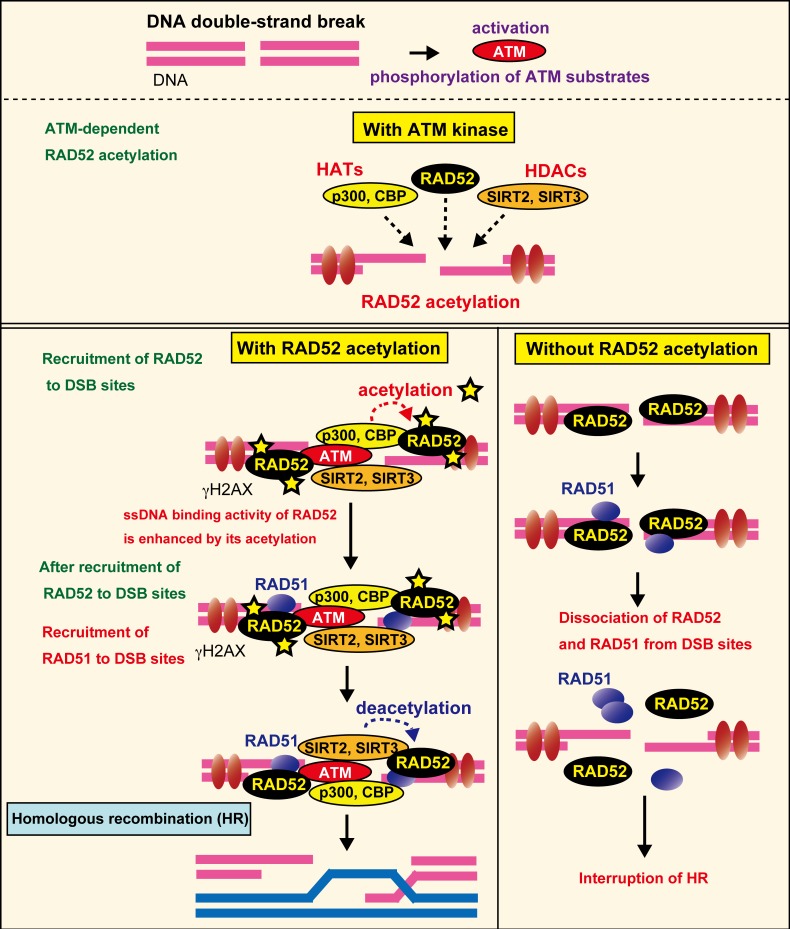
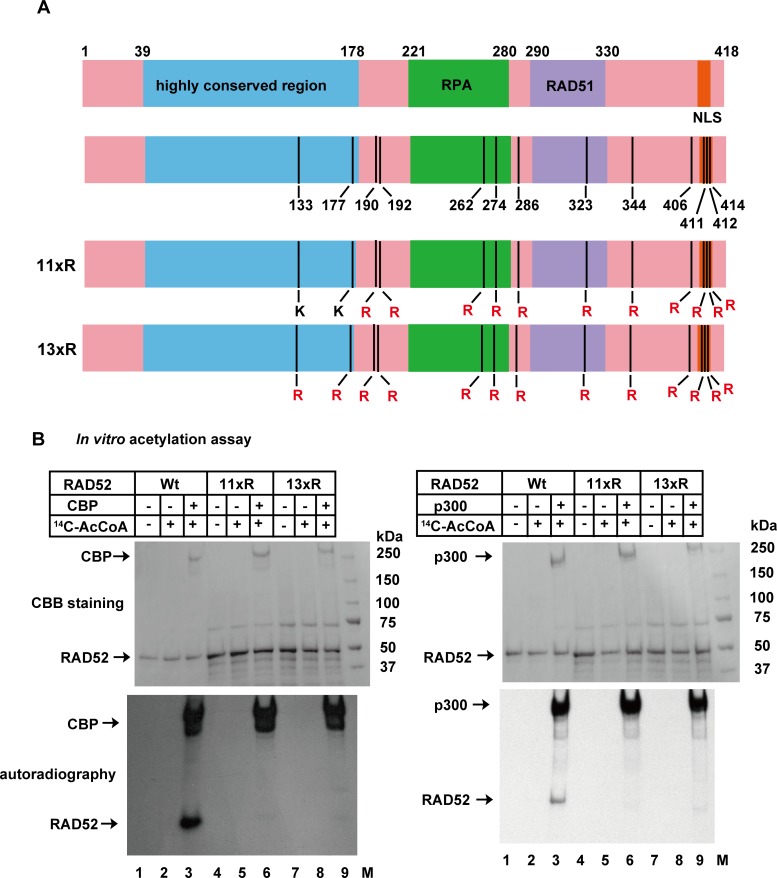
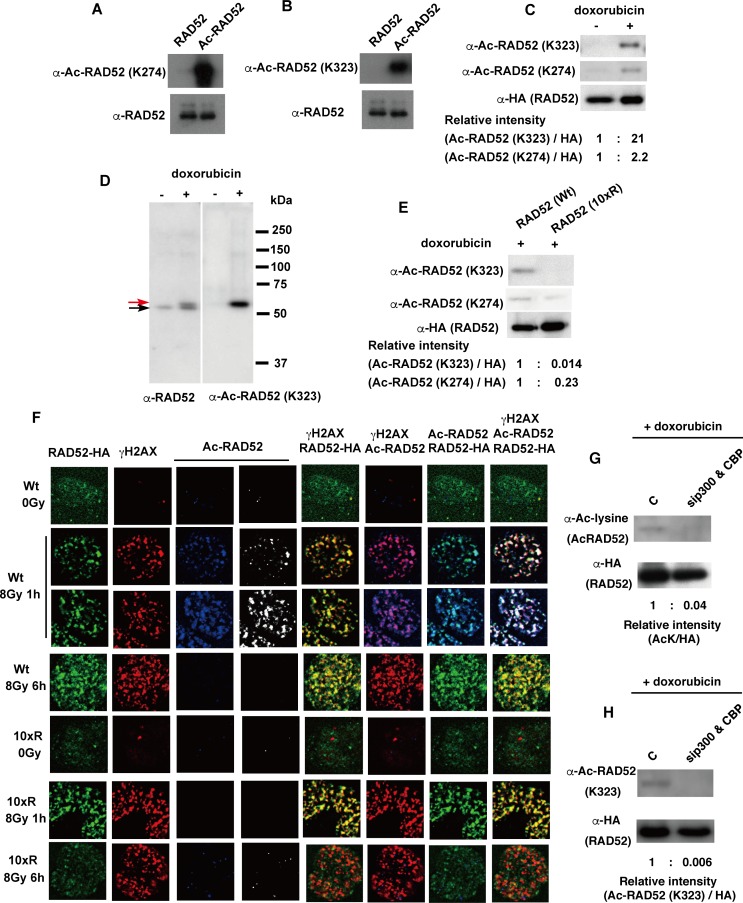
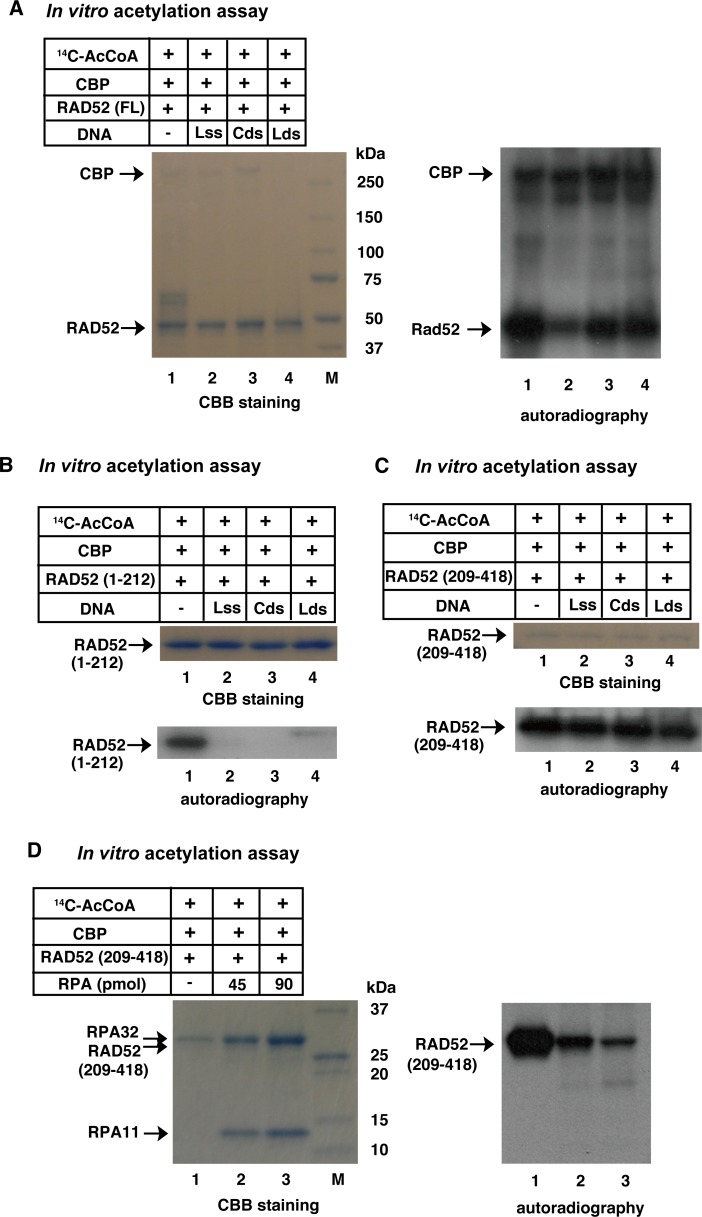
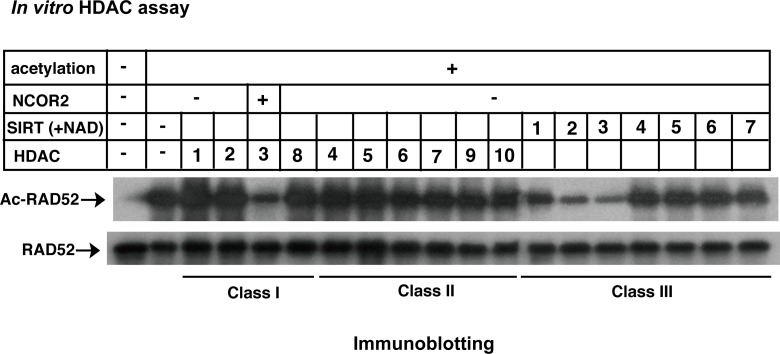
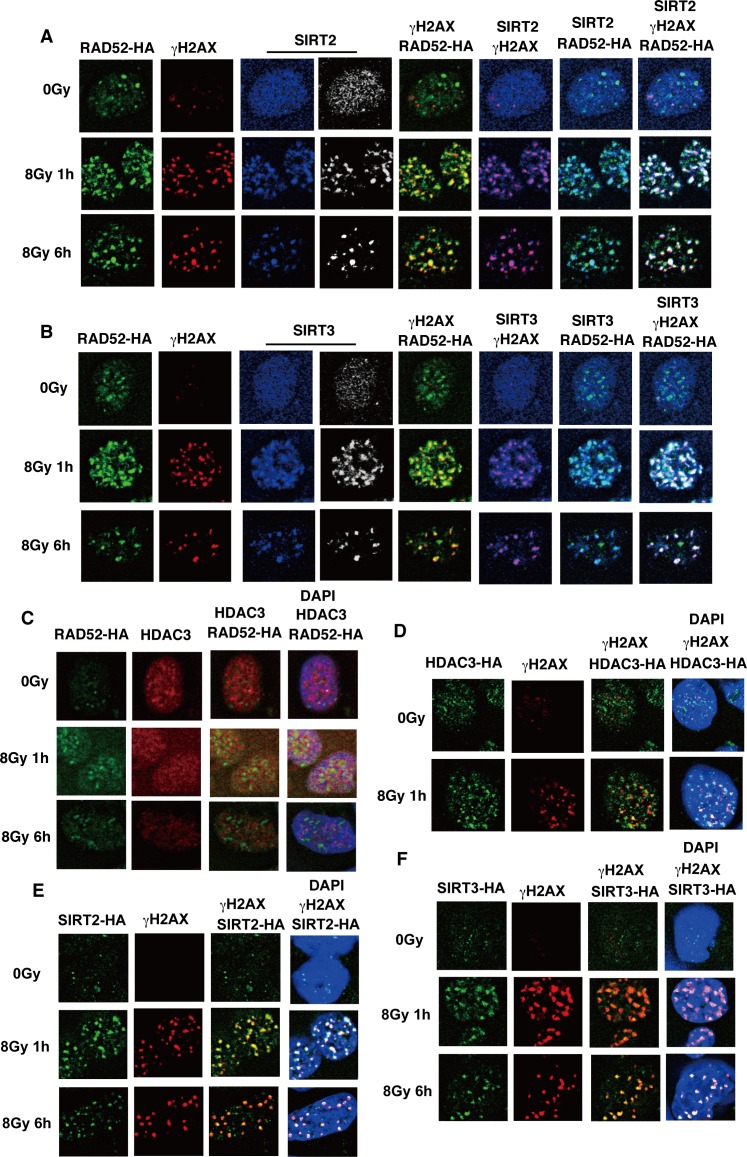
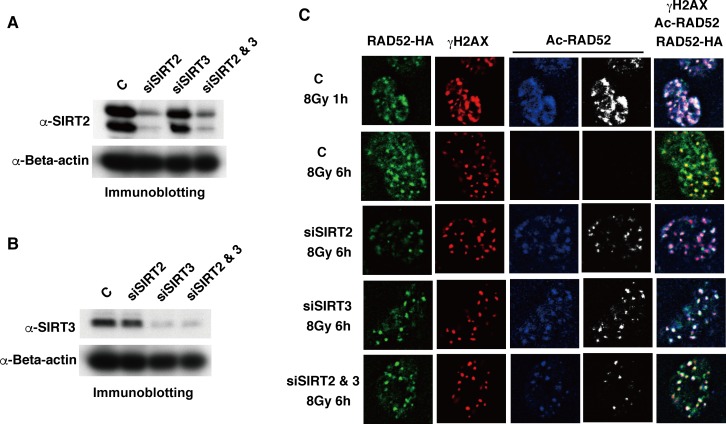
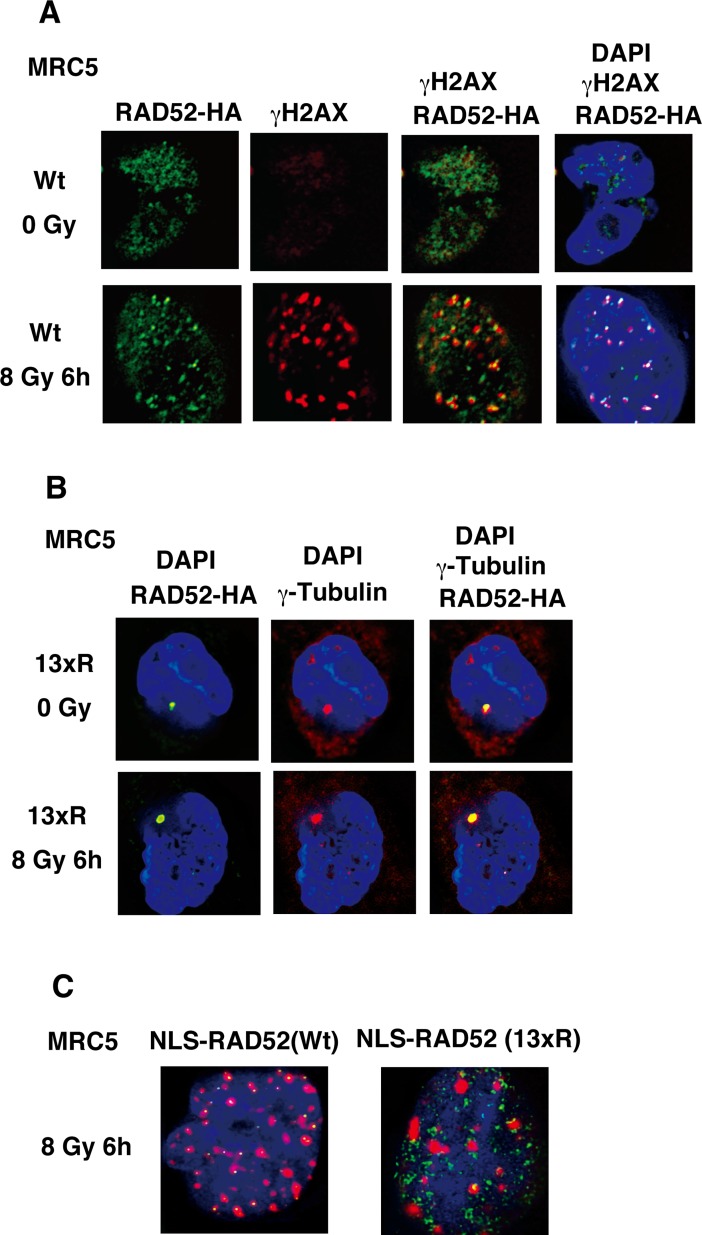
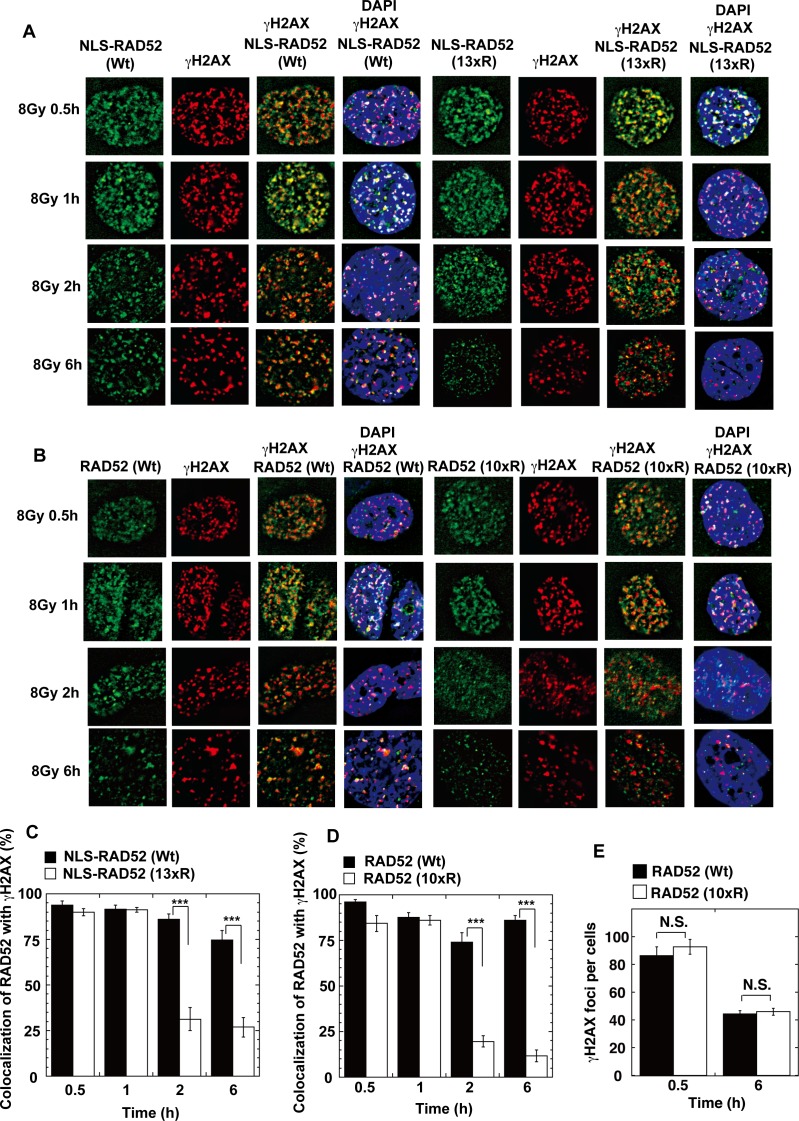
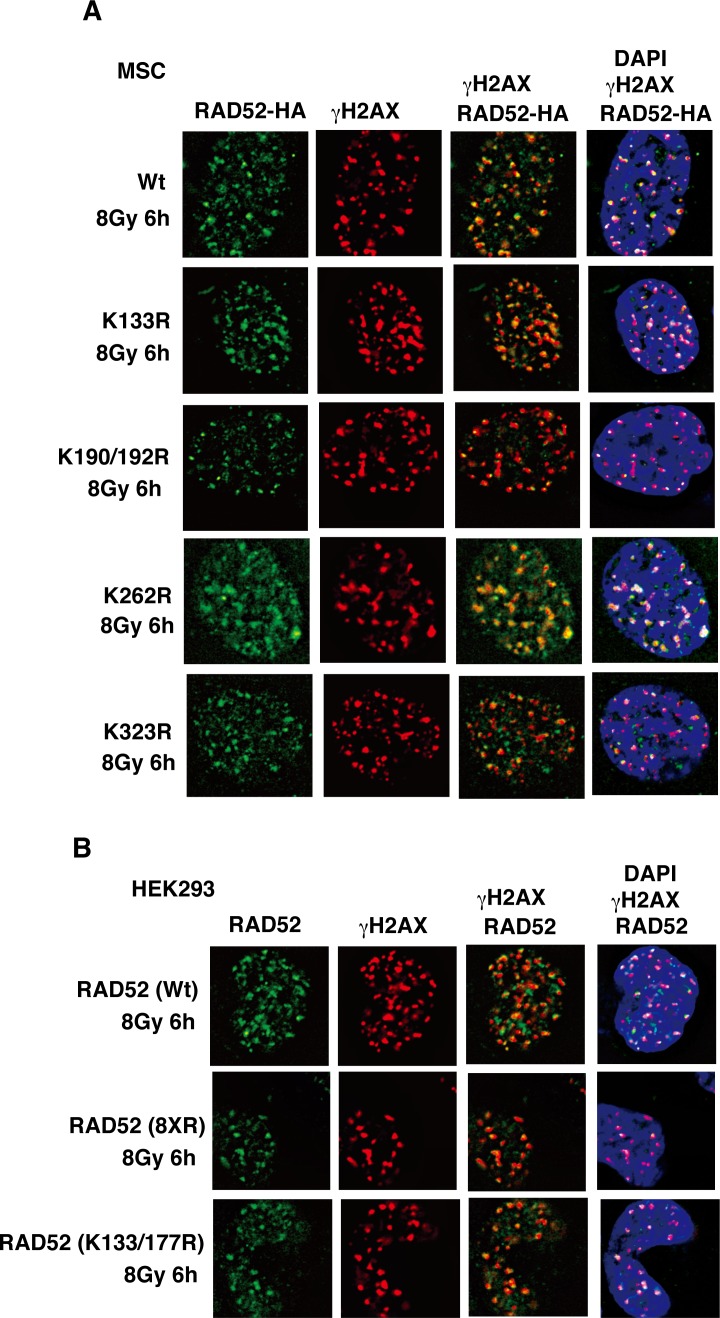
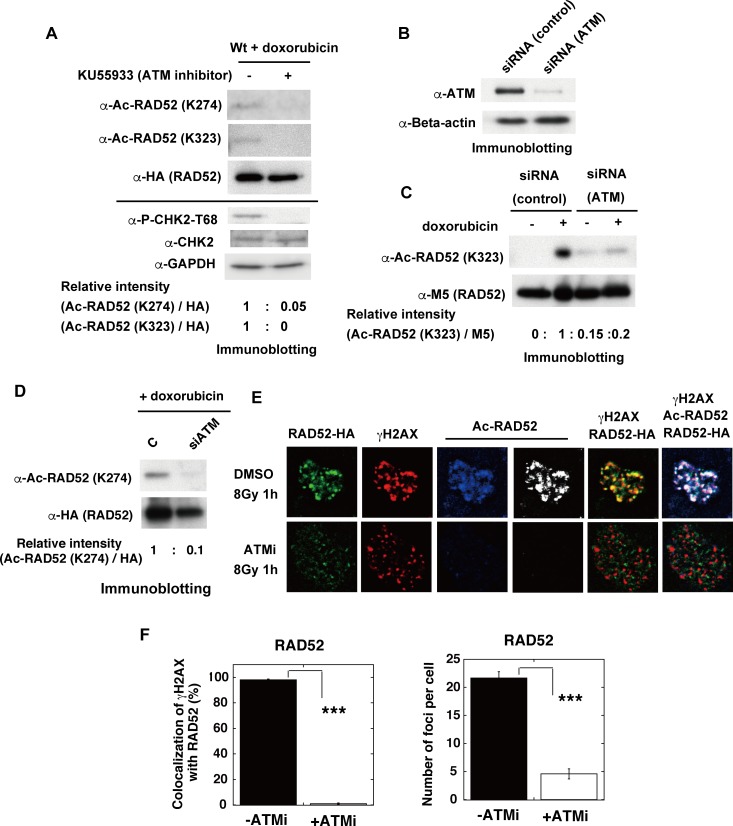
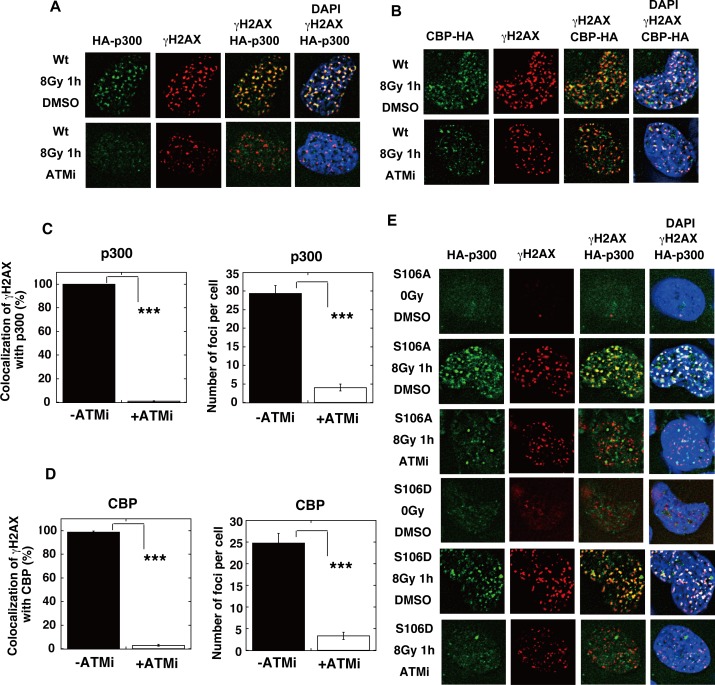
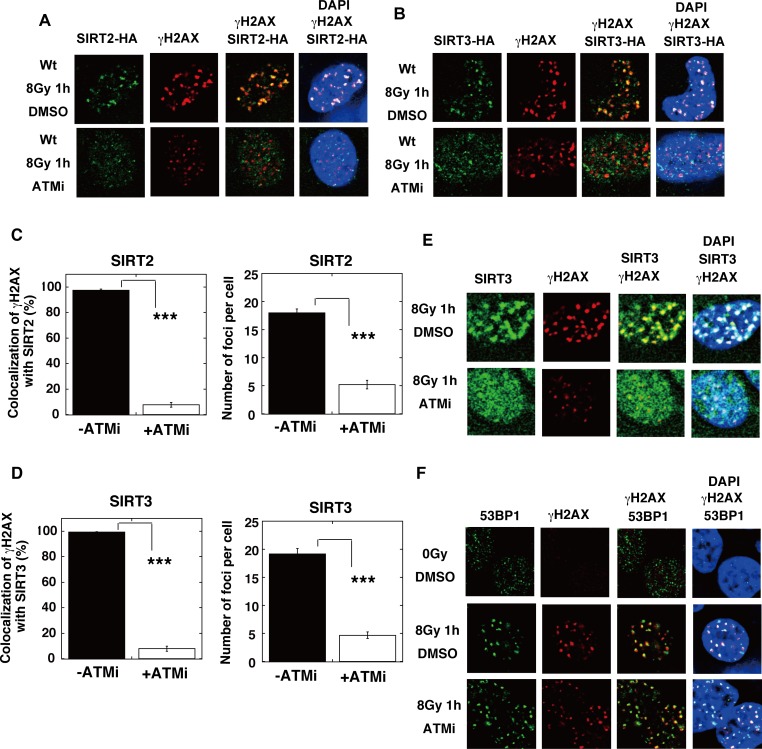
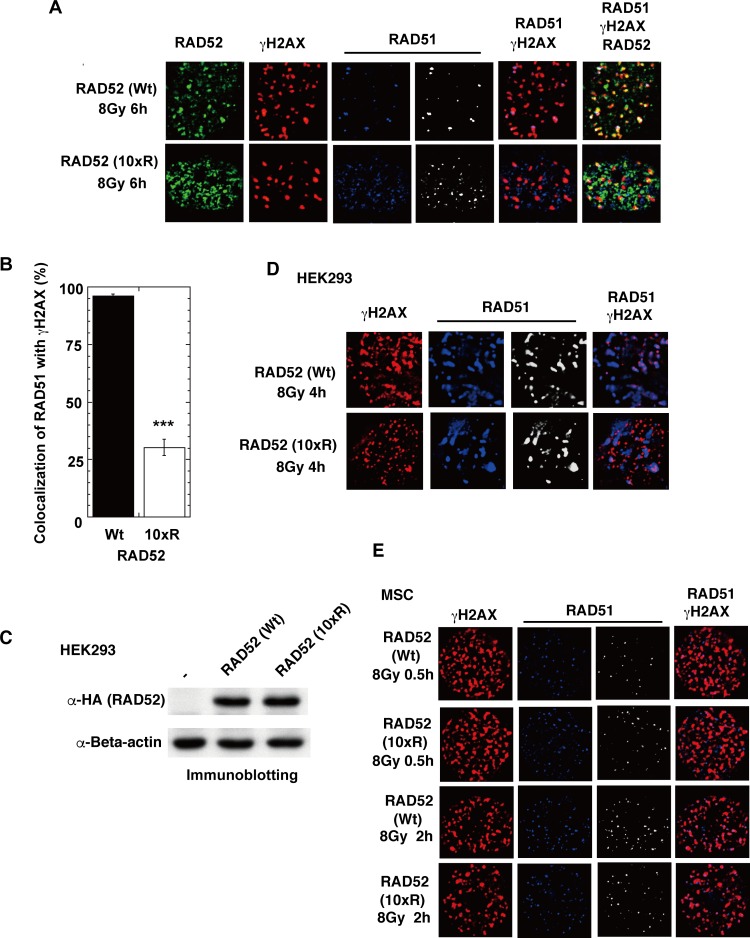
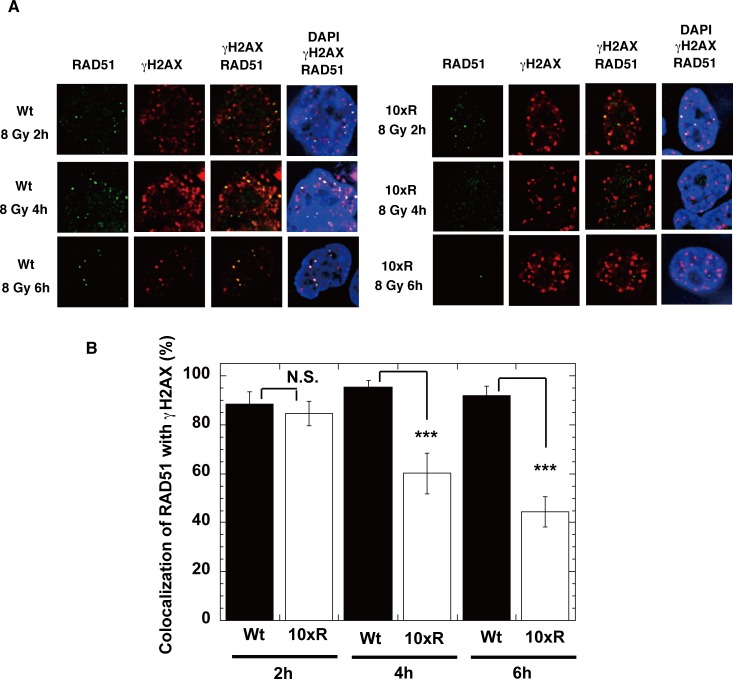
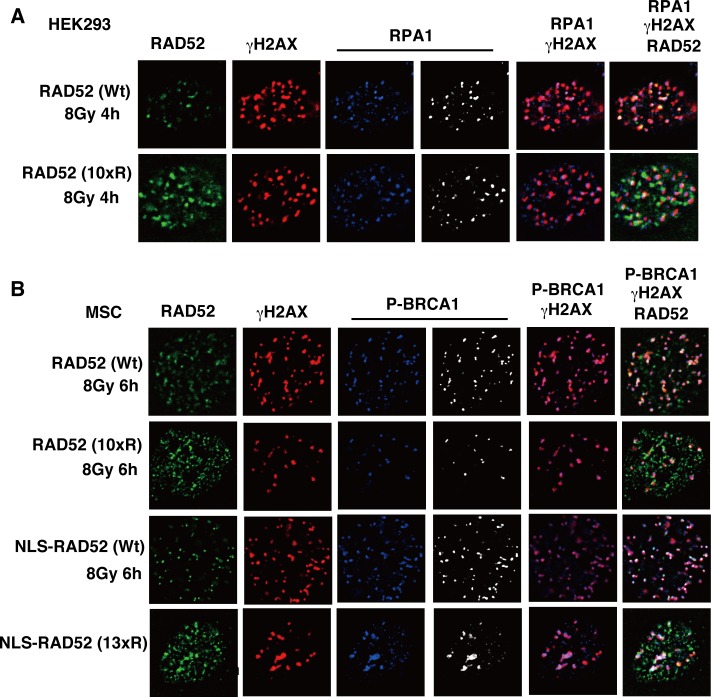
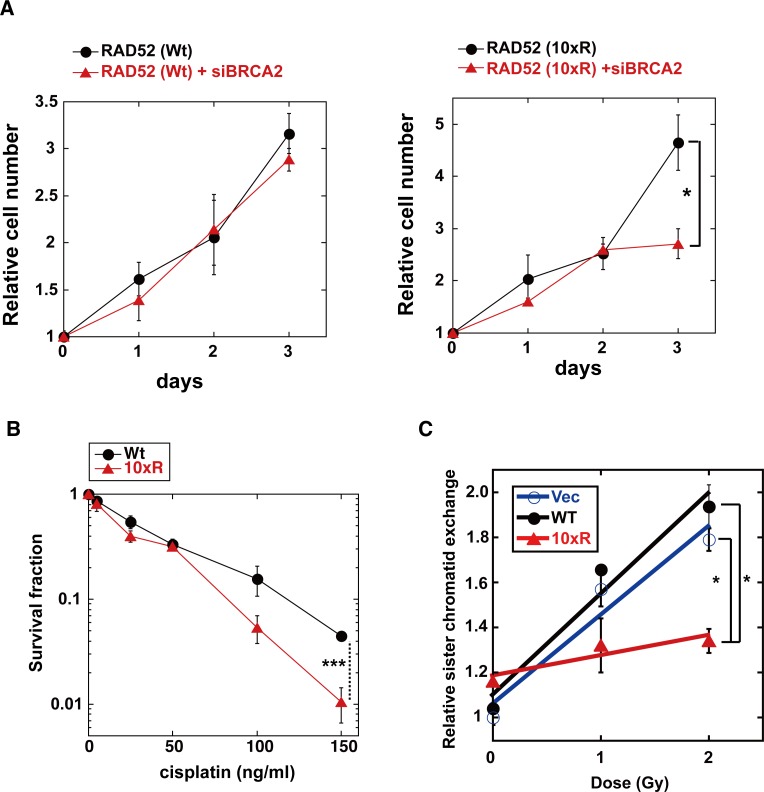
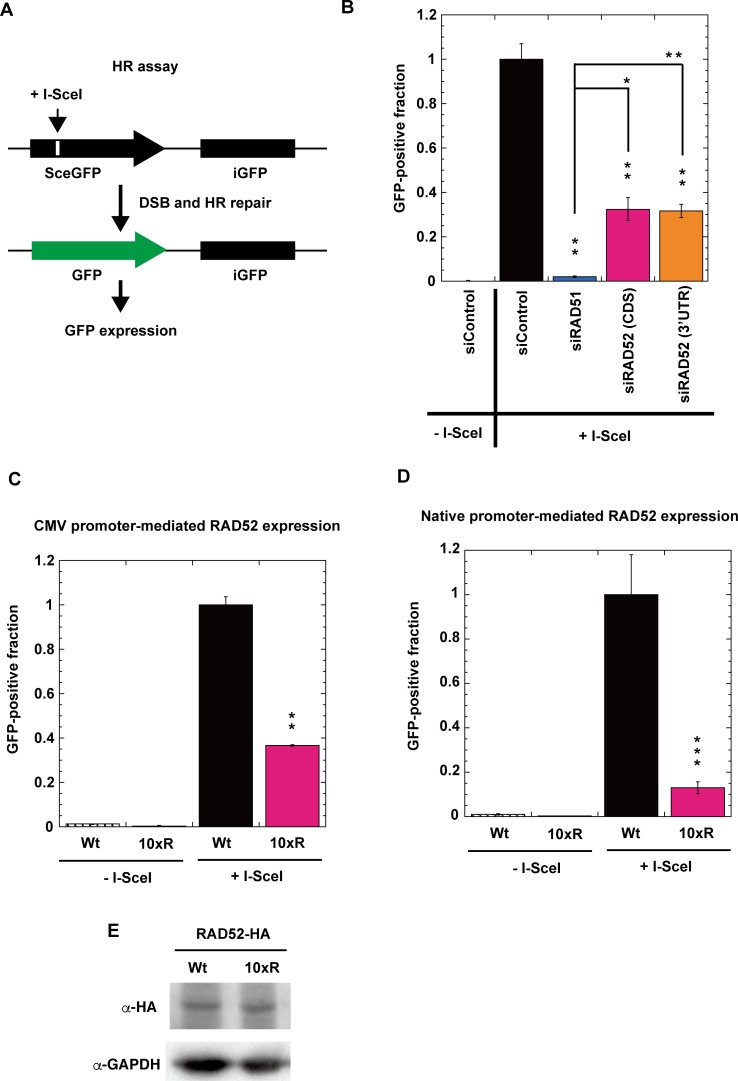
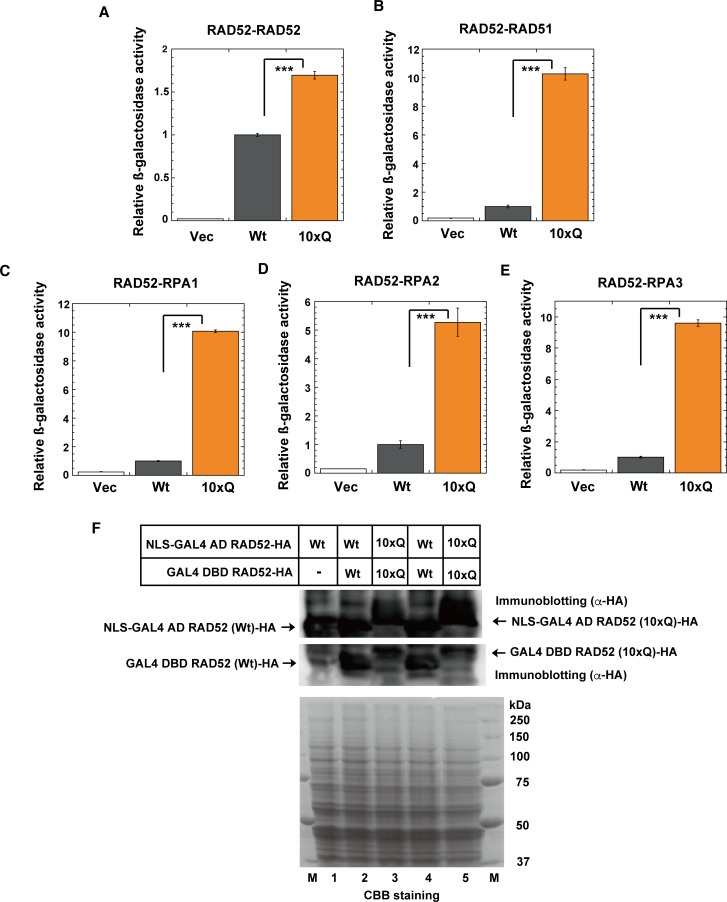
The p300 and CBP histone acetyltransferases are recruited to DNA double-strand break (DSB) sites where they induce histone acetylation, thereby influencing the chromatin structure and DNA repair process. Whether p300/CBP at DSB sites also acetylate non-histone proteins, and how their acetylation affects DSB repair, remain unknown. Here we show that p300/CBP acetylate RAD52, a human homologous recombination (HR) DNA repair protein, at DSB sites. Using in vitro acetylated RAD52, we identified 13 potential acetylation sites in RAD52 by a mass spectrometry analysis. An immunofluorescence microscopy analysis revealed that RAD52 acetylation at DSBs sites is counteracted by SIRT2- and SIRT3-mediated deacetylation, and that non-acetylated RAD52 initially accumulates at DSB sites, but dissociates prematurely from them. In the absence of RAD52 acetylation, RAD51, which plays a central role in HR, also dissociates prematurely from DSB sites, and hence HR is impaired. Furthermore, inhibition of ataxia telangiectasia mutated (ATM) protein by siRNA or inhibitor treatment demonstrated that the acetylation of RAD52 at DSB sites is dependent on the ATM protein kinase activity, through the formation of RAD52, p300/CBP, SIRT2, and SIRT3 foci at DSB sites. Our findings clarify the importance of RAD52 acetylation in HR and its underlying mechanism.
p300 和 CBP 组蛋白乙酰转移酶被招募到 DNA 双链断裂 (DSB) 位点,在那里它们诱导组蛋白乙酰化,从而影响染色质结构和 DNA 修复过程。p300/CBP 在 DSB 位点是否也乙酰化非组蛋白,以及它们的乙酰化如何影响 DSB 修复,目前尚不清楚。在这里,我们表明 p300/CBP 在 DSB 位点乙酰化 RAD52,一种人类同源重组 (HR) DNA 修复蛋白。使用体外乙酰化 RAD52,我们通过质谱分析鉴定出 RAD52 中的 13 个潜在乙酰化位点。免疫荧光显微镜分析显示,RAD52 在 DSB 位点的乙酰化受到 SIRT2 和 SIRT3 介导的去乙酰化的拮抗,并且非乙酰化的 RAD52 最初在 DSB 位点积累,但过早地从它们中解离。在 RAD52 乙酰化缺失的情况下,在 HR 中起核心作用的 RAD51 也过早地从 DSB 位点解离,因此 HR 受损。此外,通过 siRNA 或抑制剂处理抑制共济失调毛细血管扩张突变 (ATM) 蛋白,表明 RAD52 在 DSB 位点的乙酰化依赖于 ATM 蛋白激酶活性,通过在 DSB 位点形成 RAD52、p300/CBP、SIRT2 和 SIRT3 焦点。我们的发现阐明了 RAD52 乙酰化在 HR 中的重要性及其潜在机制。