Department of Biological Sciences, Faculty of Sciences, Simon Fraser University, Burnaby, BC, Canada.
Department of Cellular and Physiological Sciences, Faculty of Medicine, University of British Columbia, Vancouver, BC, Canada.
mBio. 2018 Apr 10;9(2):e02259-17. doi: 10.1128/mBio.02259-17.
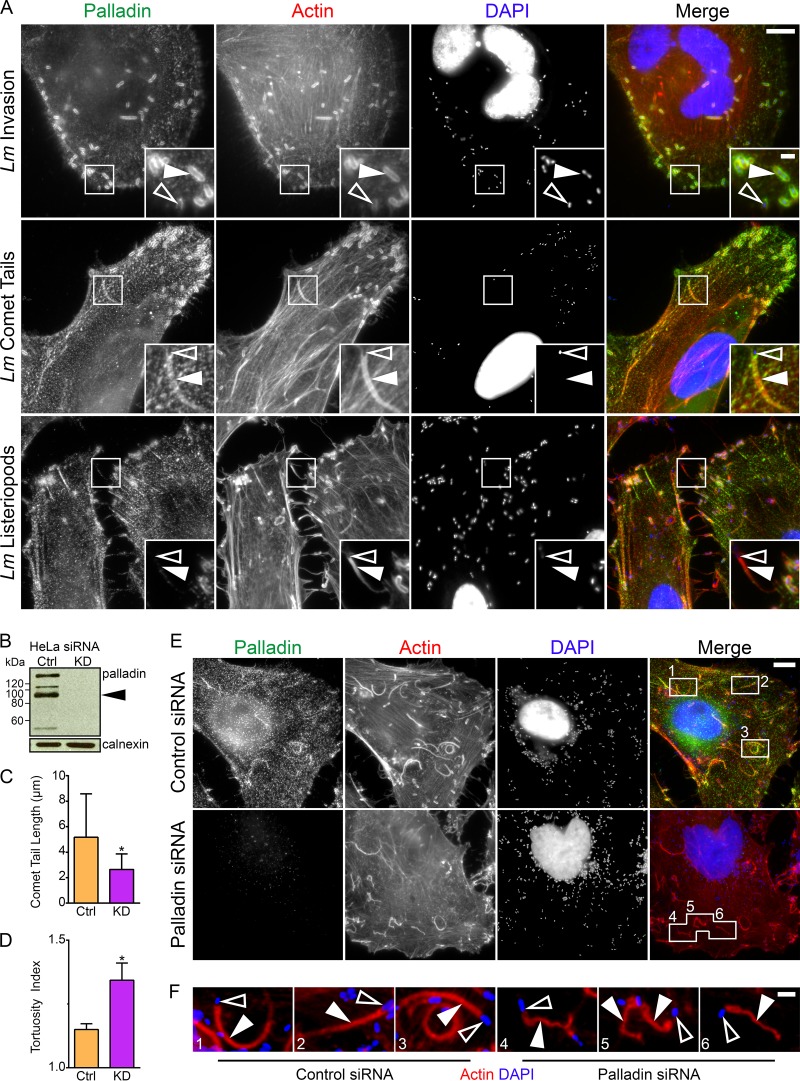
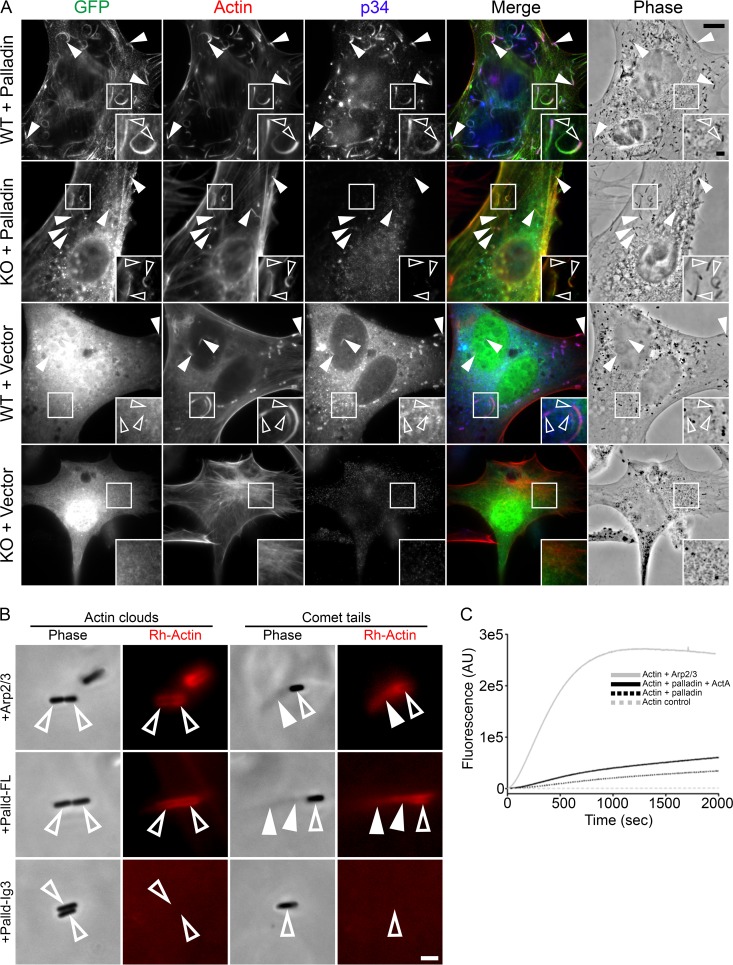
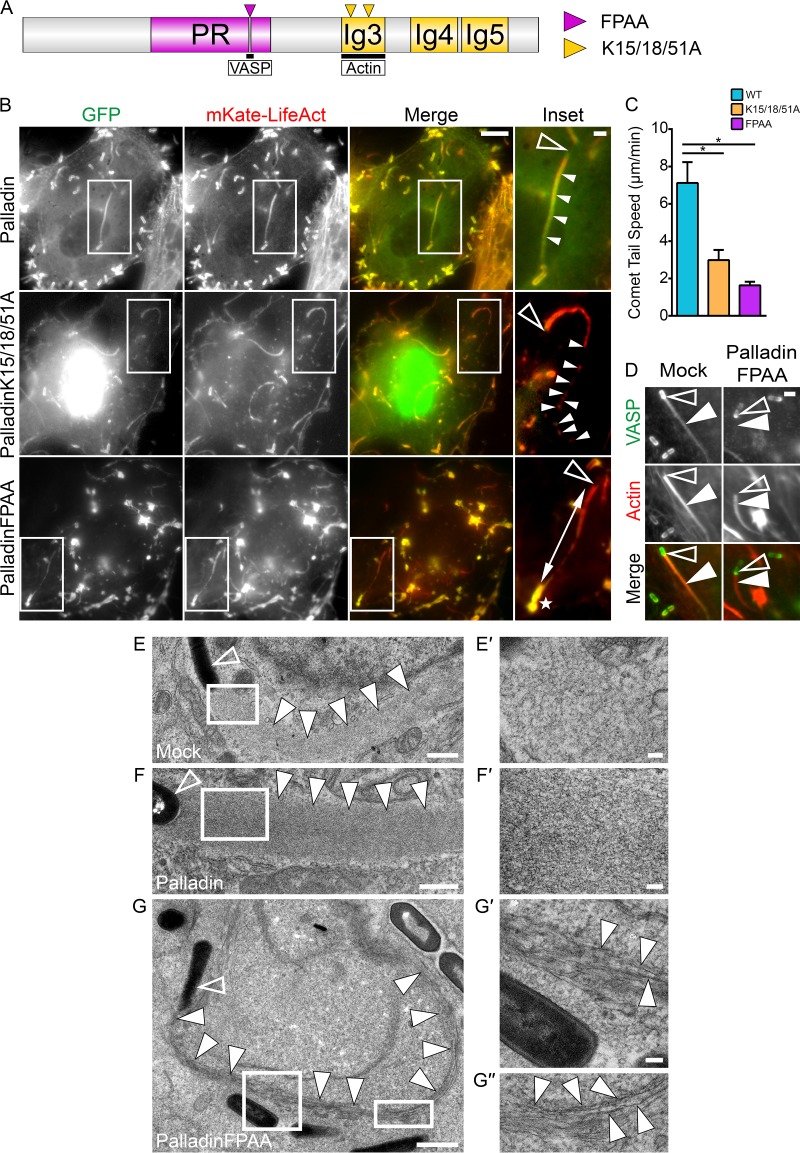
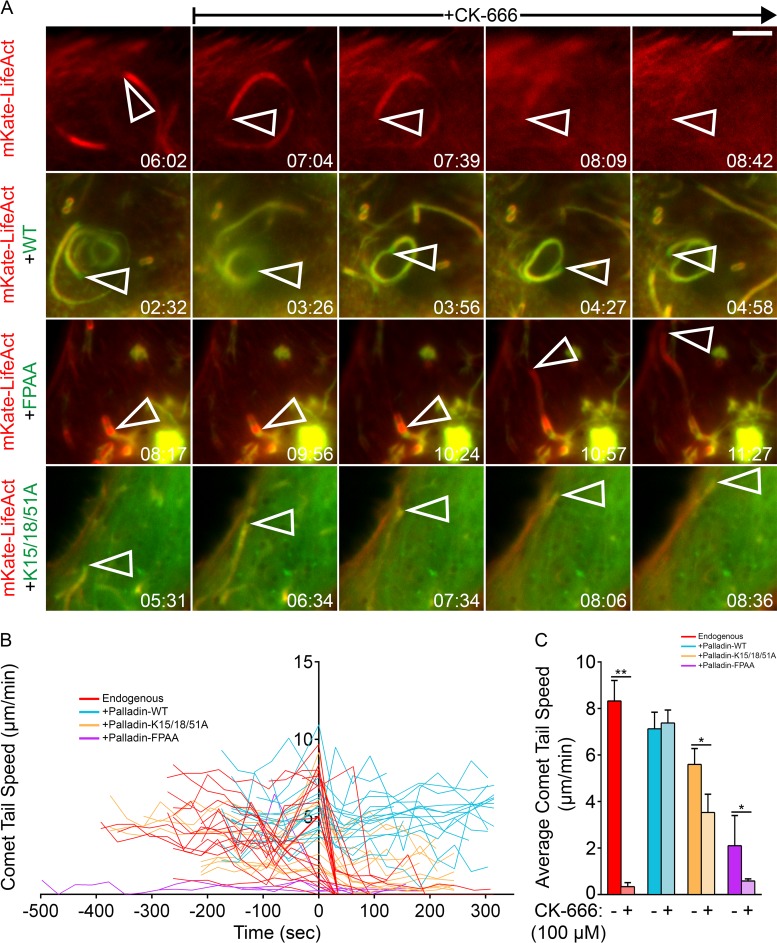
Palladin is an important component of motile actin-rich structures and nucleates branched actin filament arrays Here we examine the role of palladin during infections in order to tease out novel functions of palladin. We show that palladin is co-opted by during its cellular entry and intracellular motility. Depletion of palladin resulted in shorter and misshapen comet tails, and when actin- or VASP-binding mutants of palladin were overexpressed in cells, comet tails disintegrated or became thinner. Comet tail thinning resulted in parallel actin bundles within the structures. To determine whether palladin could compensate for the Arp2/3 complex, we overexpressed palladin in cells treated with the Arp2/3 inhibitor CK-666. In treated cells, bacterial motility could be initiated and maintained when levels of palladin were increased. To confirm these findings, we utilized a cell line depleted of multiple Arp2/3 complex subunits. Within these cells, failed to generate comet tails. When palladin was overexpressed in this Arp2/3 functionally null cell line, the ability of to generate comet tails was restored. Using purified protein components, we demonstrate that actin clouds and comet tails can be generated (in a cell-free system) by palladin in the absence of the Arp2/3 complex. Collectively, our results demonstrate that palladin can functionally replace the Arp2/3 complex during bacterial actin-based motility. Structures containing branched actin filaments require the Arp2/3 complex. One of the most commonly used systems to study intracellular movement generated by Arp2/3-based actin motility exploits actin-rich comet tails made by Using these infections together with live imaging and cell-free protein reconstitution experiments, we show that another protein, palladin, can be used in place of Arp2/3 to form actin-rich structures. Additionally, we show that palladin is needed for the structural integrity of comet tails as its depletion or mutation of critical regions causes dramatic changes to comet tail organization. These findings are the first to identify a protein that can functionally replace the Arp2/3 complex and have implications for all actin-based structures thought to exclusively use that complex.
帕拉丁是运动性富含肌动蛋白结构的重要组成部分,并起始分支肌动蛋白丝阵列。在这里,我们研究了帕拉丁在 感染期间的作用,以梳理出帕拉丁的新功能。我们表明,帕拉丁在 细胞进入和细胞内运动期间被 劫持。帕拉丁的耗竭导致彗星尾变短和变形,并且当细胞中转染帕拉丁的肌动蛋白或 VASP 结合突变体时,彗星尾瓦解或变细。彗星尾变细导致结构内的平行肌动蛋白束。为了确定帕拉丁是否可以补偿 Arp2/3 复合物,我们在用 Arp2/3 抑制剂 CK-666 处理的细胞中转染帕拉丁。在处理的细胞中,当帕拉丁水平增加时,可以启动和维持细菌运动。为了证实这些发现,我们利用耗尽多个 Arp2/3 复合物亚基的细胞系。在这些细胞中, 不能生成彗星尾。当帕拉丁在这个 Arp2/3 功能缺失的细胞系中转染时, 生成彗星尾的能力得到恢复。使用纯化的蛋白成分,我们证明,在没有 Arp2/3 复合物的情况下,帕拉丁可以在无细胞系统中生成 actin 云团和彗星尾。总之,我们的结果表明,帕拉丁可以在细菌肌动蛋白依赖的运动中替代 Arp2/3 复合物发挥功能。包含分支肌动蛋白丝的结构需要 Arp2/3 复合物。研究基于 Arp2/3 肌动蛋白运动的细胞内运动的最常用系统之一是利用 actin 丰富的彗星尾,这些彗星尾由 生成。使用这些感染以及活细胞成像和无细胞蛋白重建实验,我们表明,另一种蛋白帕拉丁可以替代 Arp2/3 形成 actin 丰富的结构。此外,我们表明,帕拉丁是彗星尾结构完整性所必需的,因为其耗竭或关键区域的突变会导致彗星尾组织的剧烈变化。这些发现是首次鉴定出一种可以替代 Arp2/3 复合物的蛋白,并对所有被认为专门使用该复合物的基于肌动蛋白的结构具有影响。