Suraneni Praveen, Fogelson Ben, Rubinstein Boris, Noguera Philippe, Volkmann Niels, Hanein Dorit, Mogilner Alex, Li Rong
Stowers Institute for Medical Research, Kansas City, MO 64110.
Courant Institute and Department of Biology, New York University, New York, NY 10012.
Mol Biol Cell. 2015 Mar 1;26(5):901-12. doi: 10.1091/mbc.E14-07-1250. Epub 2015 Jan 7.
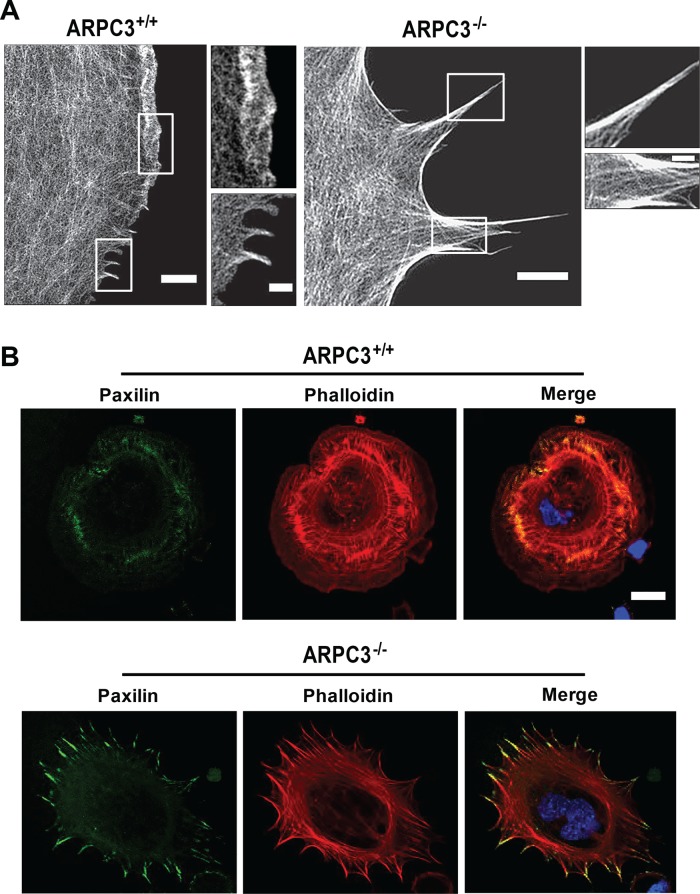
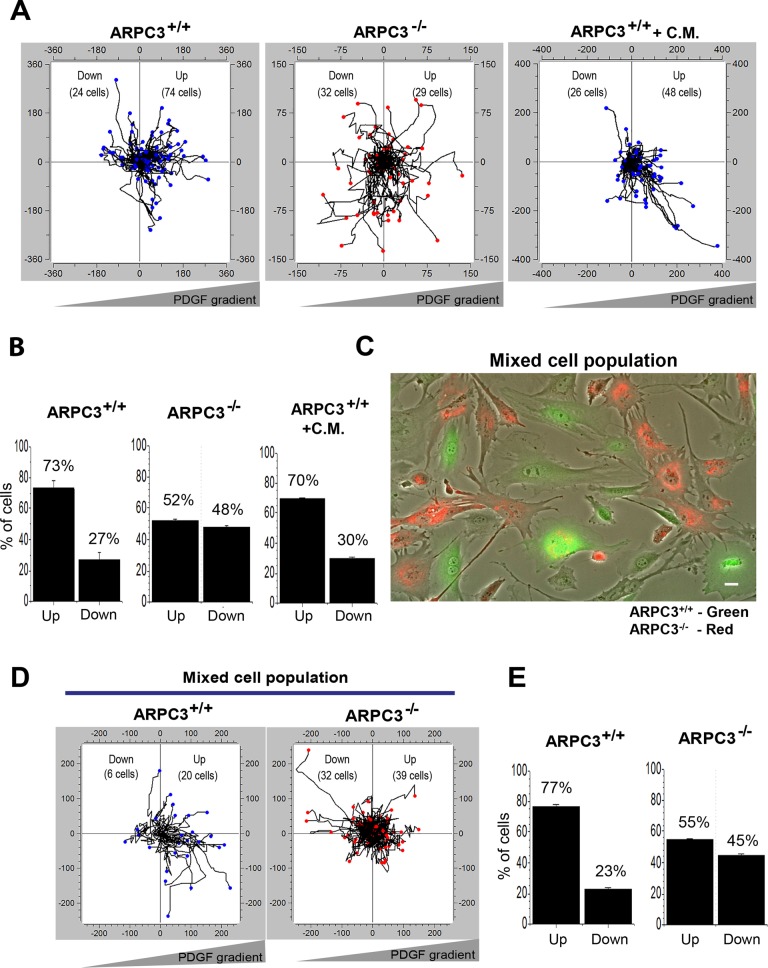
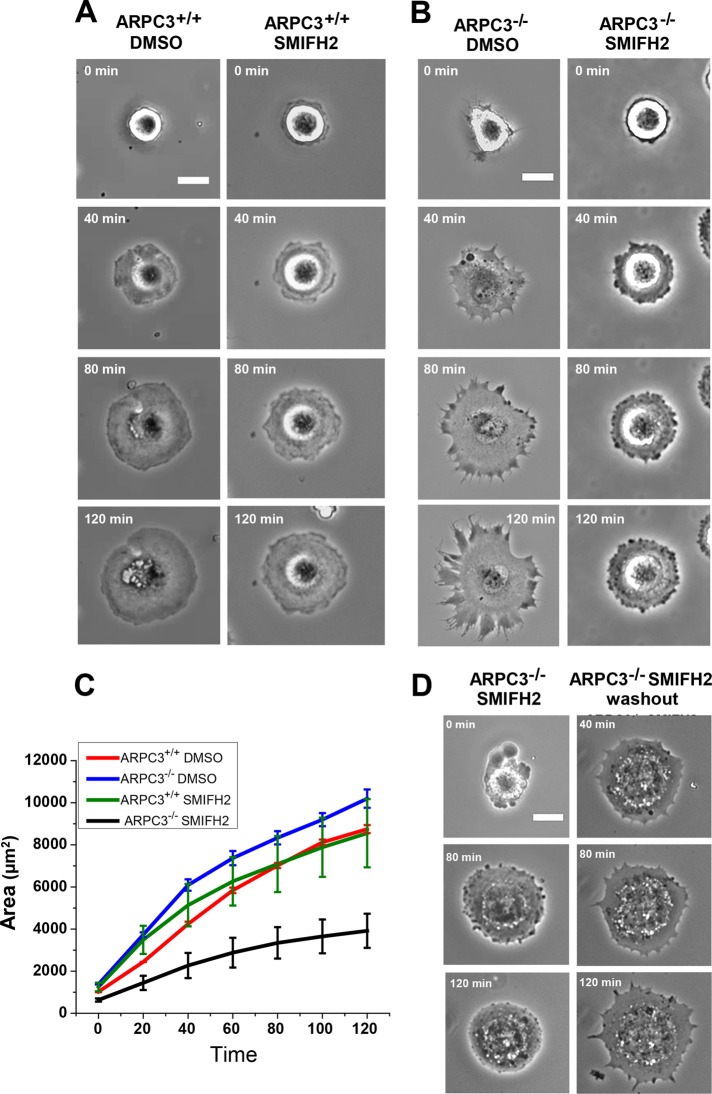
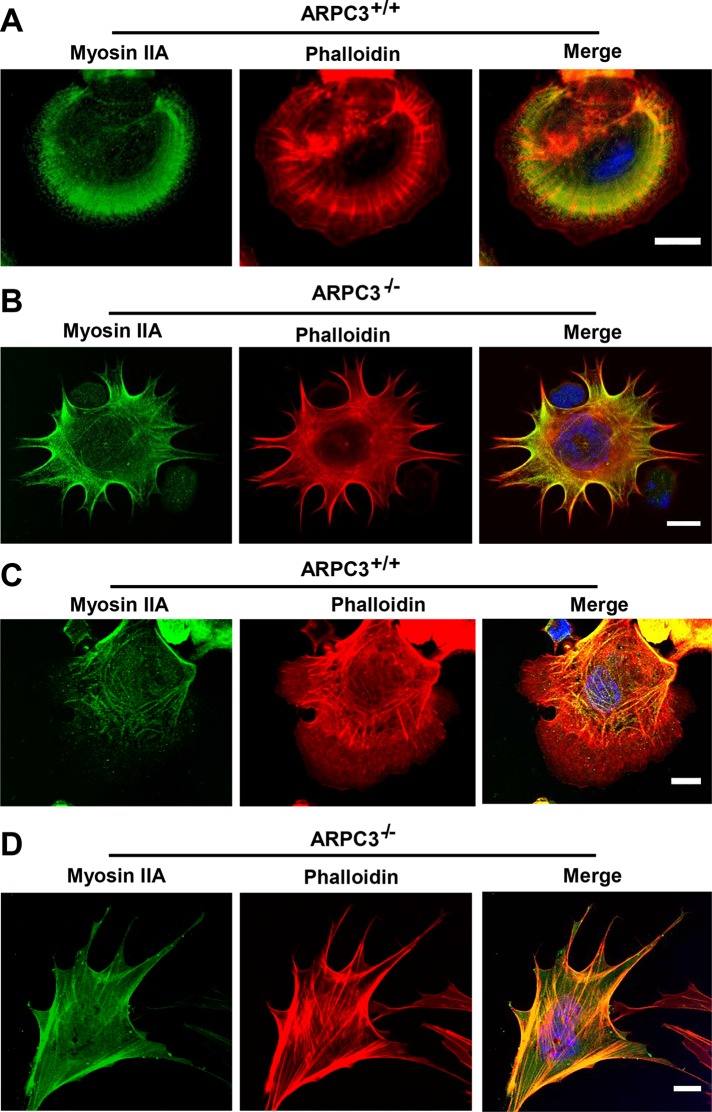
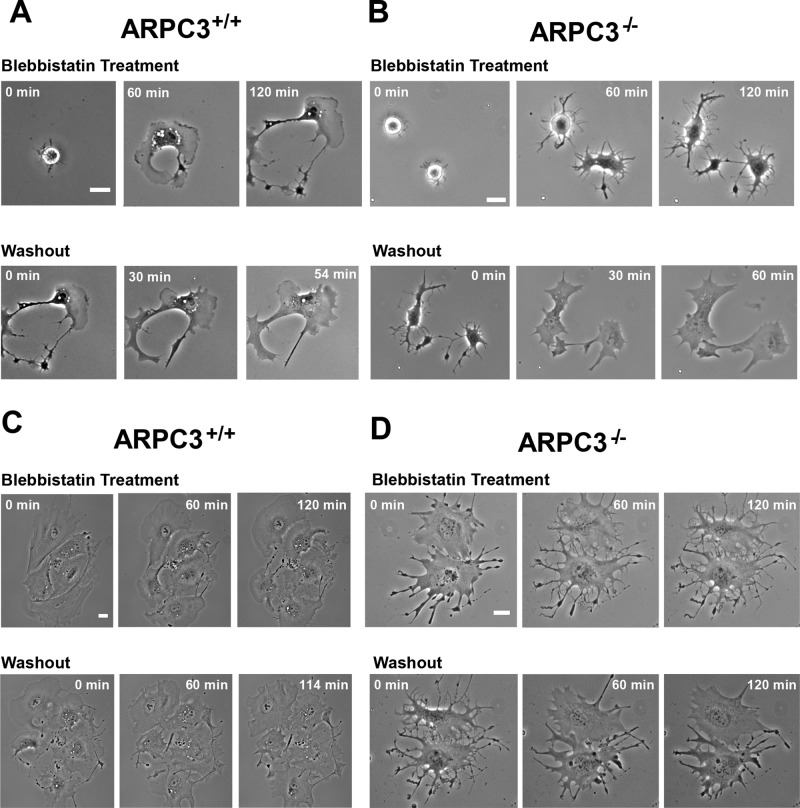
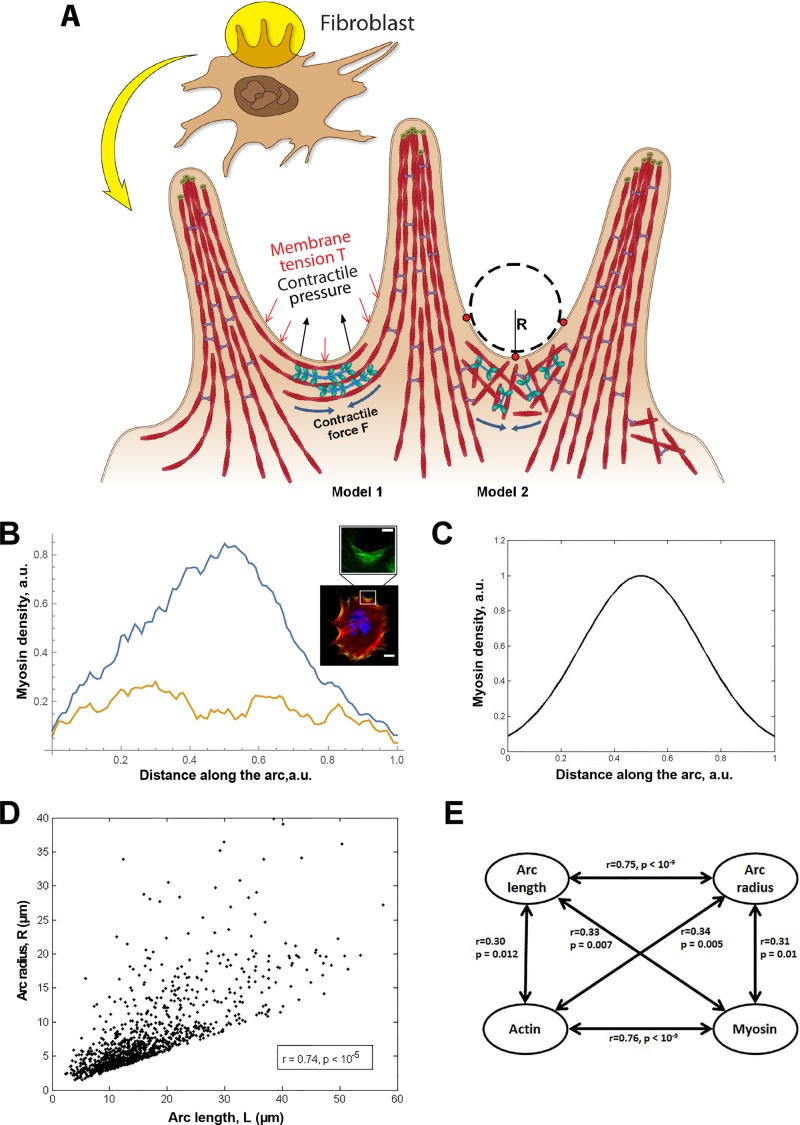
Cells employ protrusive leading edges to navigate and promote their migration in diverse physiological environments. Classical models of leading-edge protrusion rely on a treadmilling dendritic actin network that undergoes continuous assembly nucleated by the Arp2/3 complex, forming ruffling lamellipodia. Recent work demonstrated, however, that, in the absence of the Arp2/3 complex, fibroblast cells adopt a leading edge with filopodia-like protrusions (FLPs) and maintain an ability to move, albeit with altered responses to different environmental signals. We show that formin-family actin nucleators are required for the extension of FLPs but are insufficient to produce a continuous leading edge in fibroblasts lacking Arp2/3 complex. Myosin II is concentrated in arc-like regions of the leading edge in between FLPs, and its activity is required for coordinated advancement of these regions with formin-generated FLPs. We propose that actomyosin contraction acting against membrane tension advances the web of arcs between FLPs. Predictions of this model are verified experimentally. The dependence of myosin II in leading-edge advancement helps explain the previously reported defect in directional movement in the Arpc3-null fibroblasts. We provide further evidence that this defect is cell autonomous during chemotaxis.
细胞利用突出的前缘在各种生理环境中导航并促进其迁移。前缘突出的经典模型依赖于一种踏车式的树突状肌动蛋白网络,该网络通过Arp2/3复合体进行连续组装成核,形成皱褶状的片状伪足。然而,最近的研究表明,在缺乏Arp2/3复合体的情况下,成纤维细胞会形成具有丝状伪足样突起(FLP)的前缘,并保持移动能力,尽管对不同环境信号的反应有所改变。我们发现,formin家族的肌动蛋白成核剂是FLP延伸所必需的,但不足以在缺乏Arp2/3复合体的成纤维细胞中产生连续的前缘。肌球蛋白II集中在FLP之间前缘的弧形区域,其活性是这些区域与formin产生的FLP协同推进所必需的。我们提出,肌动球蛋白收缩对抗膜张力推动了FLP之间的弧形网络前进。该模型的预测通过实验得到了验证。肌球蛋白II在前缘推进中的依赖性有助于解释先前报道的Arpc3基因敲除成纤维细胞定向运动缺陷。我们提供了进一步的证据表明,这种缺陷在趋化作用中是细胞自主性的。