Department of Physics and Astronomy, University of Denver, Denver, United States.
Department of Biological Sciences, University of Denver, Denver, United States.
Elife. 2018 Jul 9;7:e34586. doi: 10.7554/eLife.34586.
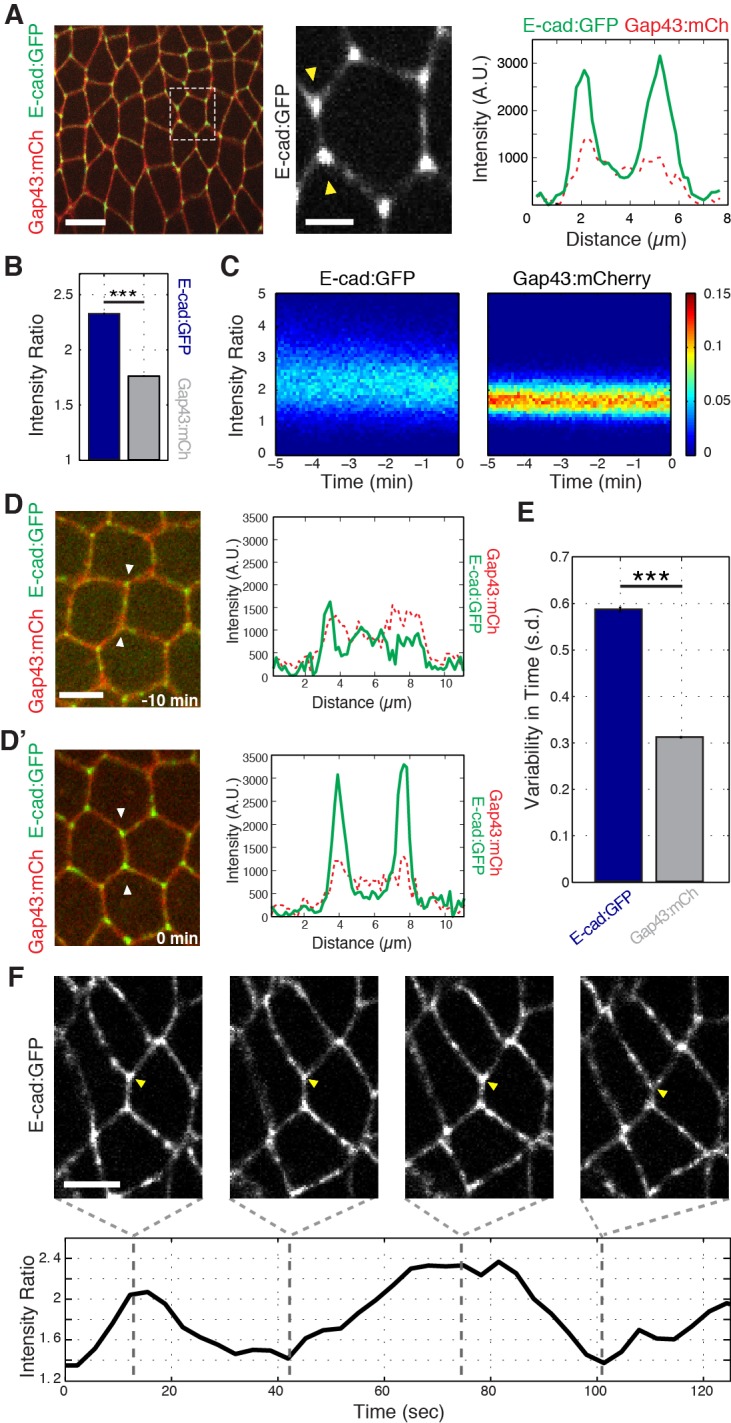
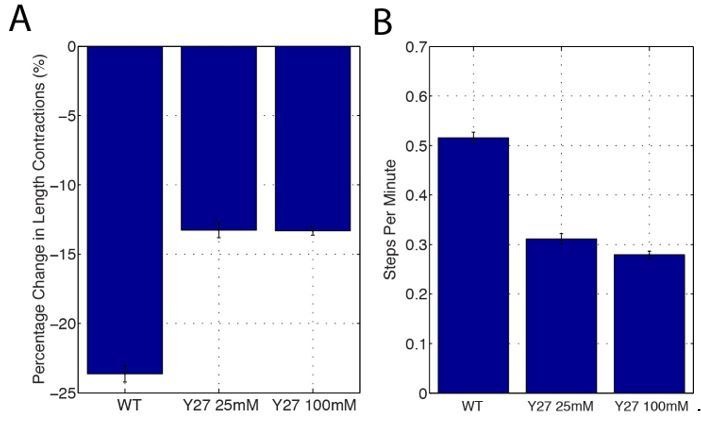
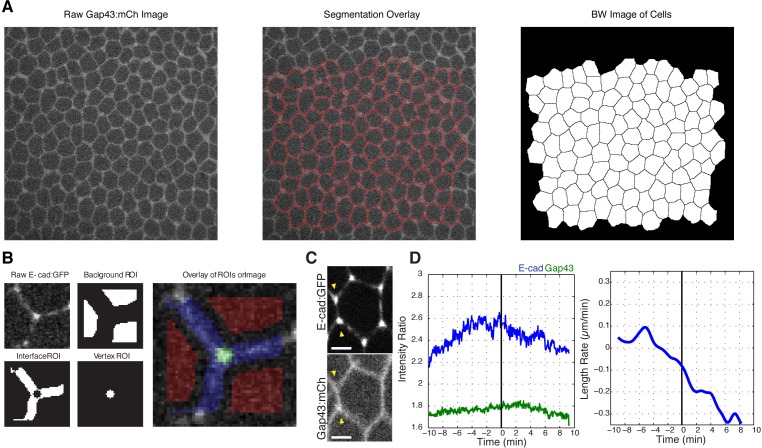
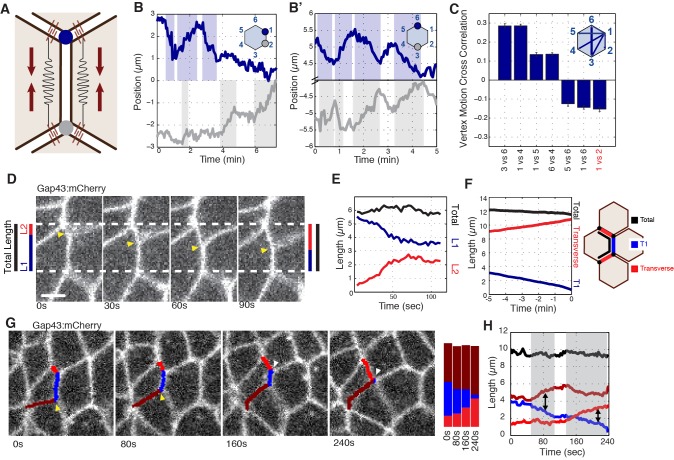
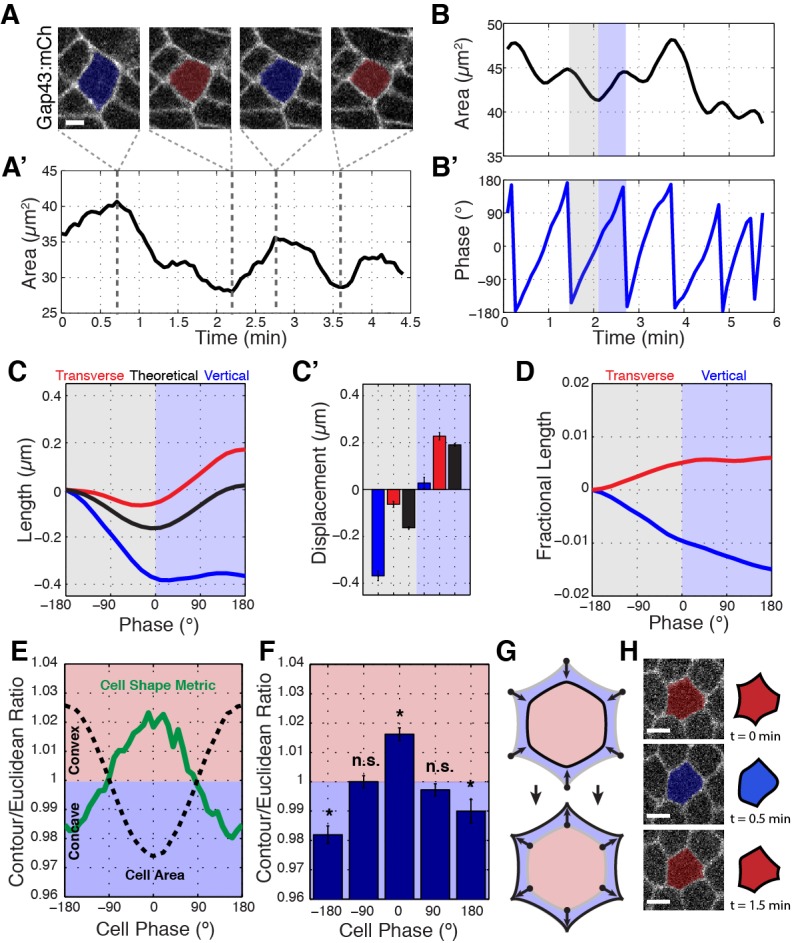
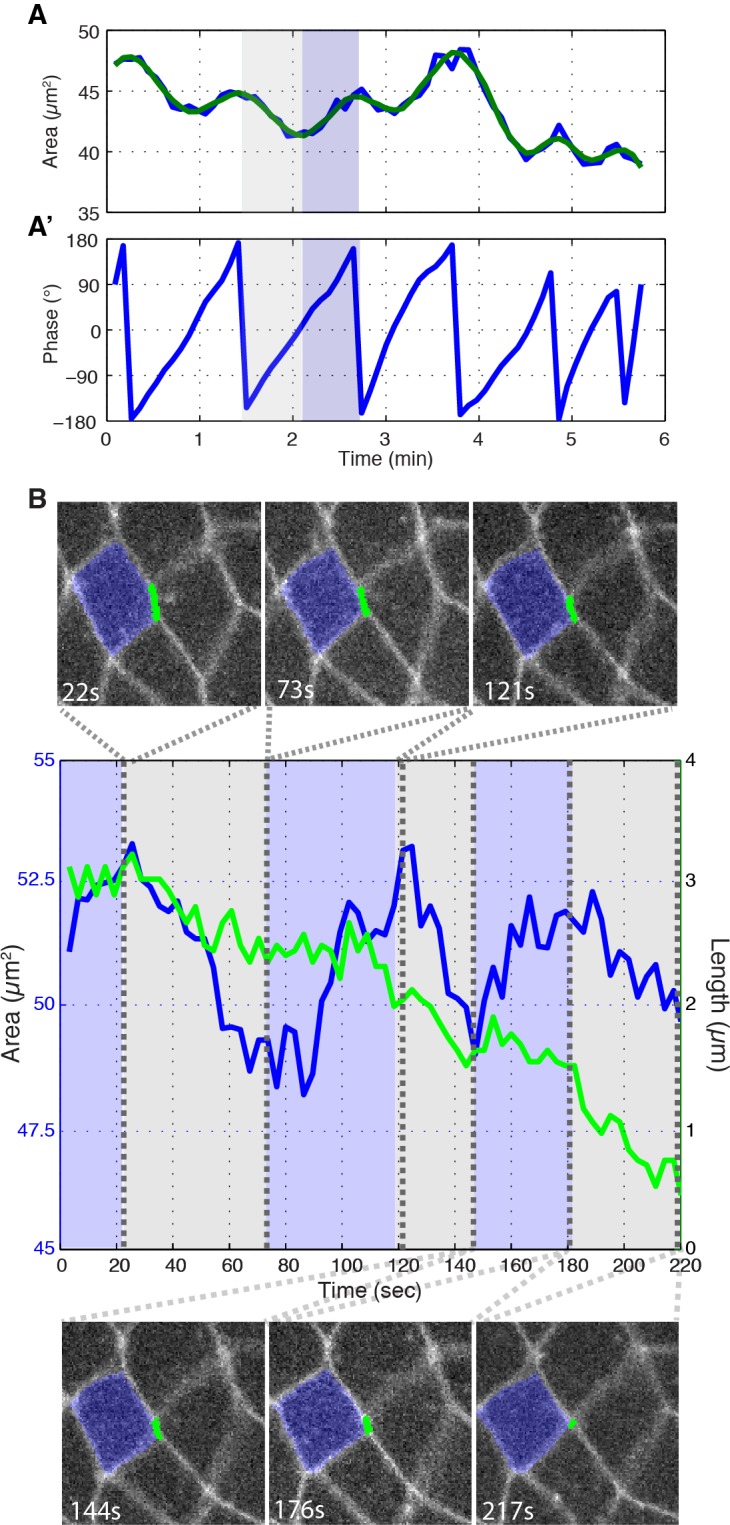
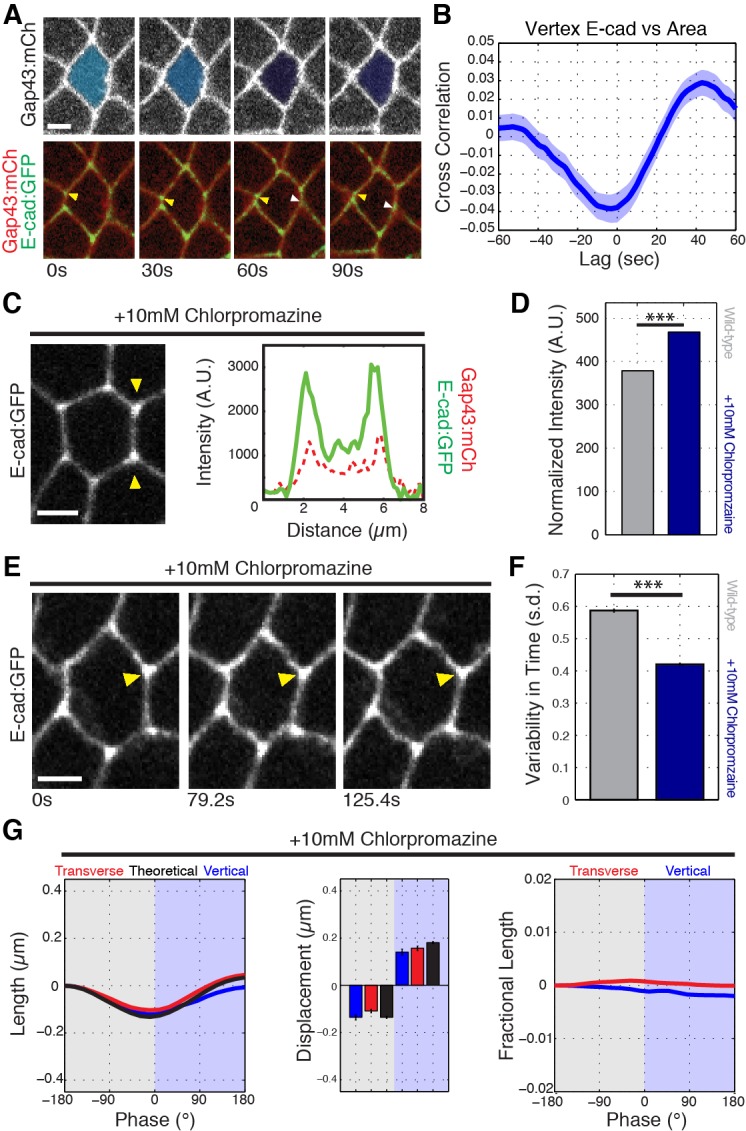
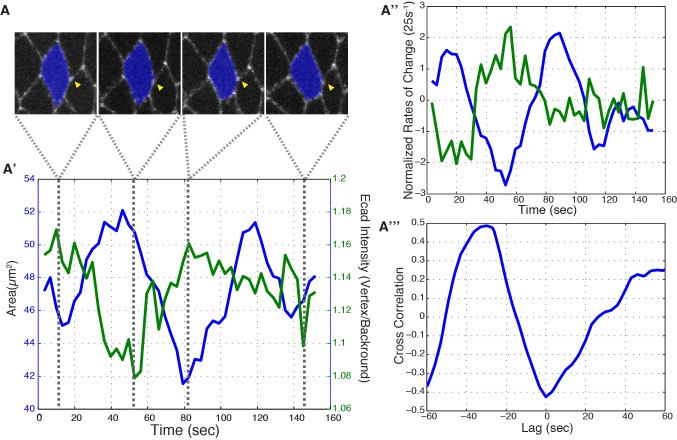
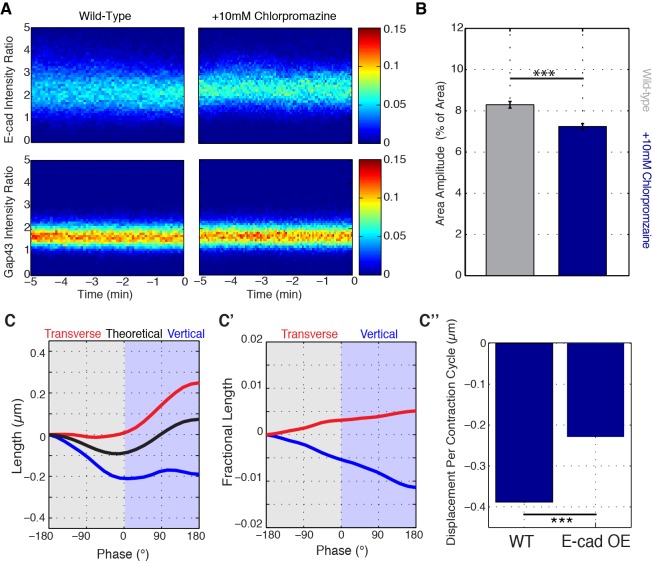
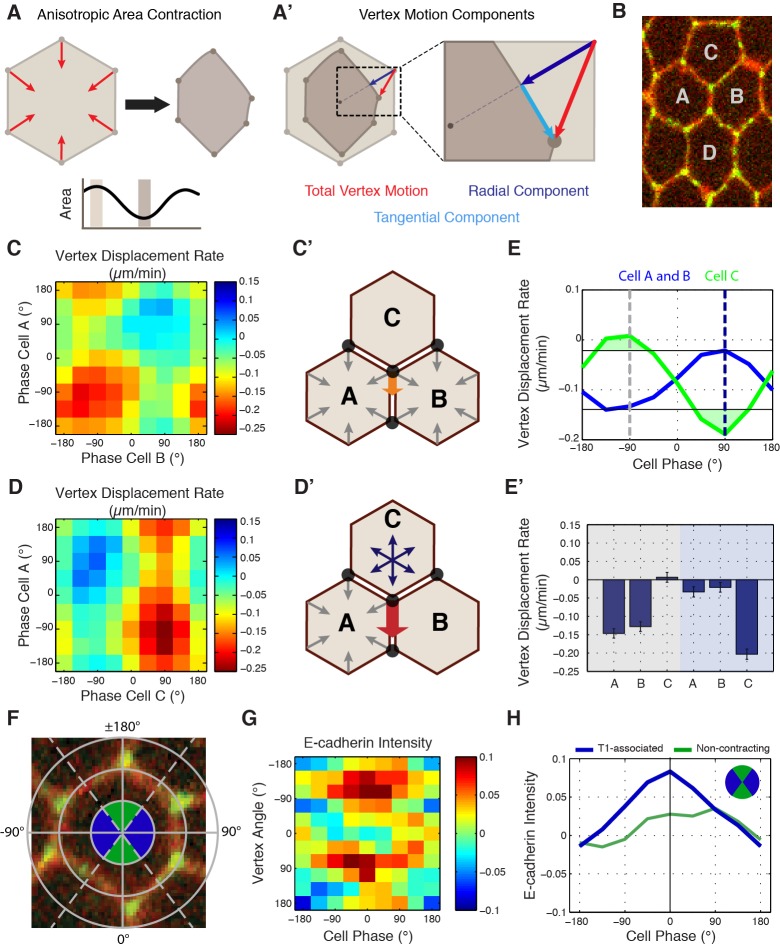
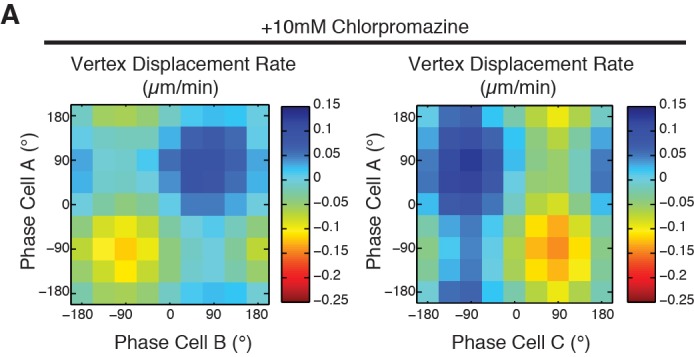
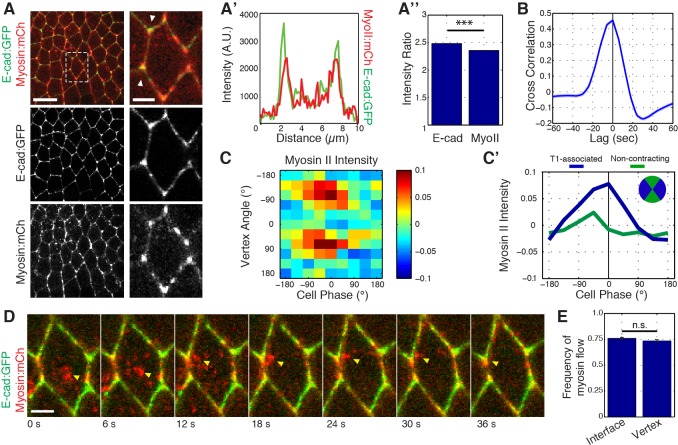
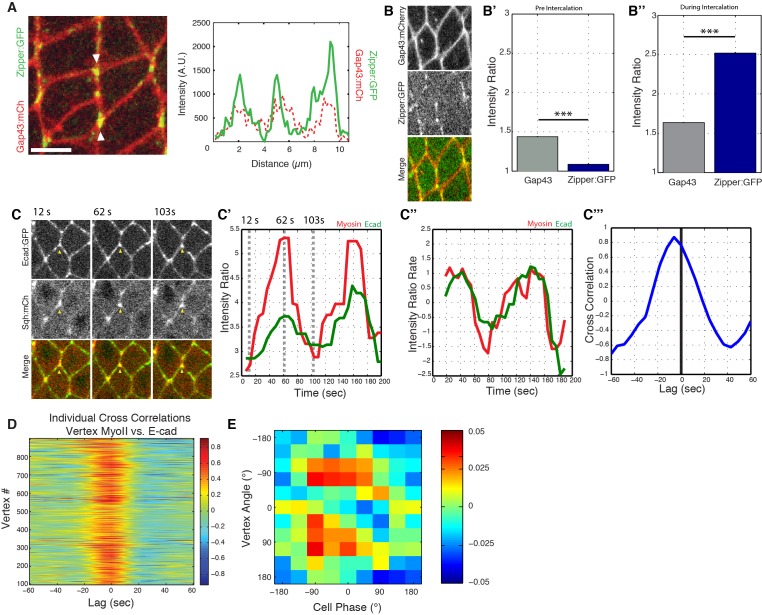
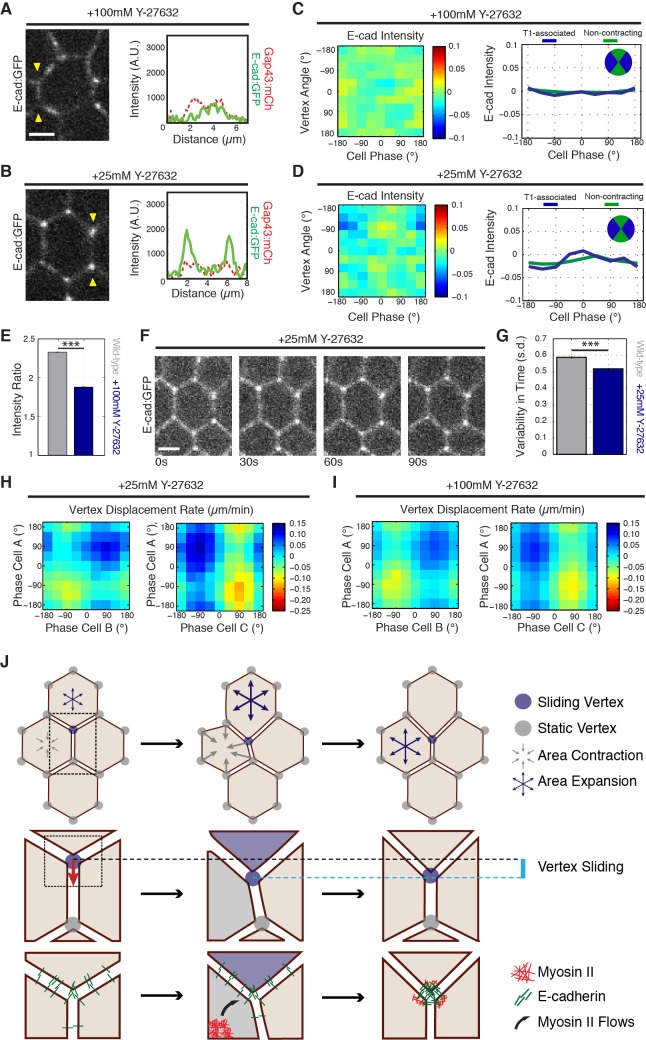
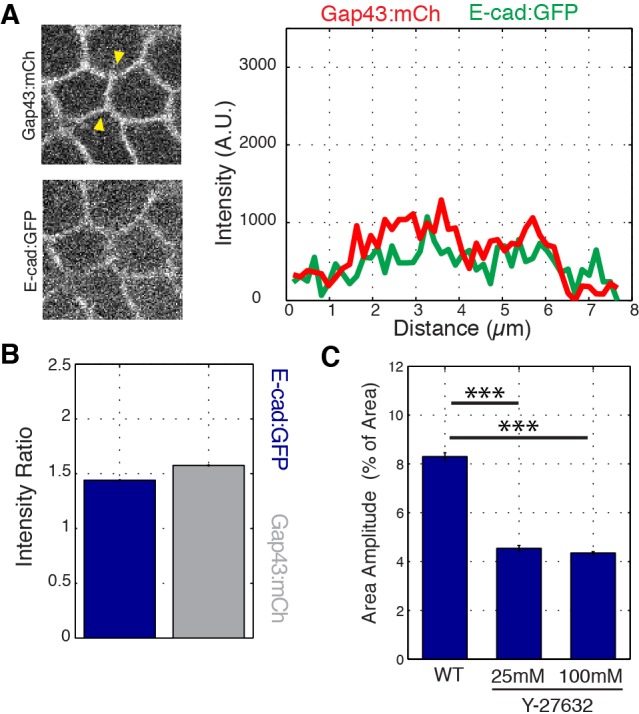
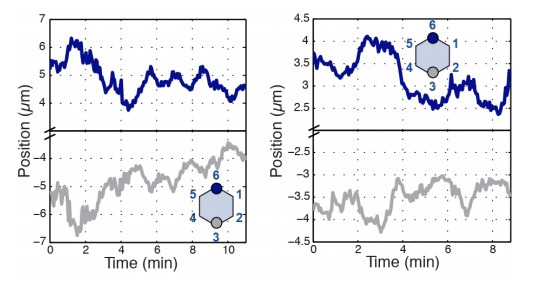
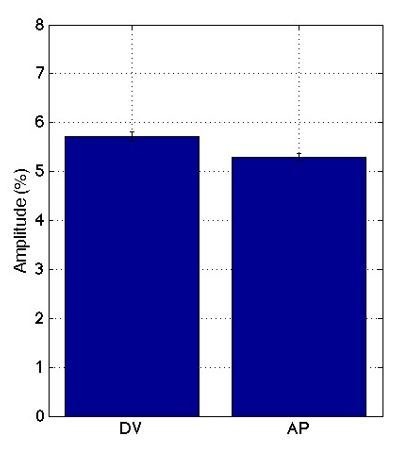
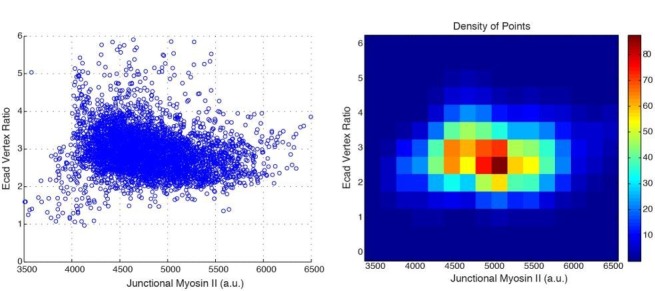
Oriented cell intercalation is an essential developmental process that shapes tissue morphologies through the directional insertion of cells between their neighbors. Previous research has focused on properties of cellcell , while the function of tricellular has remained unaddressed. Here, we identify a highly novel mechanism in which vertices demonstrate independent sliding behaviors along cell peripheries to produce the topological deformations responsible for intercalation. Through systematic analysis, we find that the motion of vertices connected by contracting interfaces is not physically coupled, but instead possess strong radial coupling. E-cadherin and Myosin II exist in previously unstudied populations at cell vertices and undergo oscillatory cycles of accumulation and dispersion that are coordinated with changes in cell area. Additionally, peak enrichment of vertex E-cadherin/Myosin II coincides with interface length stabilization. Our results suggest a model in which asymmetric radial force balance directs the progressive, ratcheted motion of individual vertices to drive intercalation.
定向细胞插入是一个基本的发育过程,通过细胞在相邻细胞之间的定向插入来塑造组织形态。以前的研究集中在细胞-细胞间的特性上,而三细胞间的作用仍然没有得到解决。在这里,我们发现了一个非常新颖的机制,即顶点沿细胞边缘表现出独立的滑动行为,产生负责插入的拓扑变形。通过系统分析,我们发现由收缩界面连接的顶点的运动在物理上没有耦合,而是具有很强的径向耦合。E-钙粘蛋白和肌球蛋白 II 存在于细胞顶点以前未研究过的群体中,并经历与细胞面积变化相协调的积累和分散的振荡循环。此外,顶点 E-钙粘蛋白/肌球蛋白 II 的峰值富集与界面长度稳定一致。我们的结果表明,一种模型认为不对称的径向力平衡指导单个顶点的渐进、棘轮式运动,从而驱动插入。