Department of Genetics, Yale School of Medicine, New Haven, CT 06520, USA.
Howard Hughes Medical Institute, Department of Molecular Biosciences, University of Texas at Austin, Austin, TX 78712, USA.
Sci Signal. 2018 Jul 10;11(538):eaaq0702. doi: 10.1126/scisignal.aaq0702.
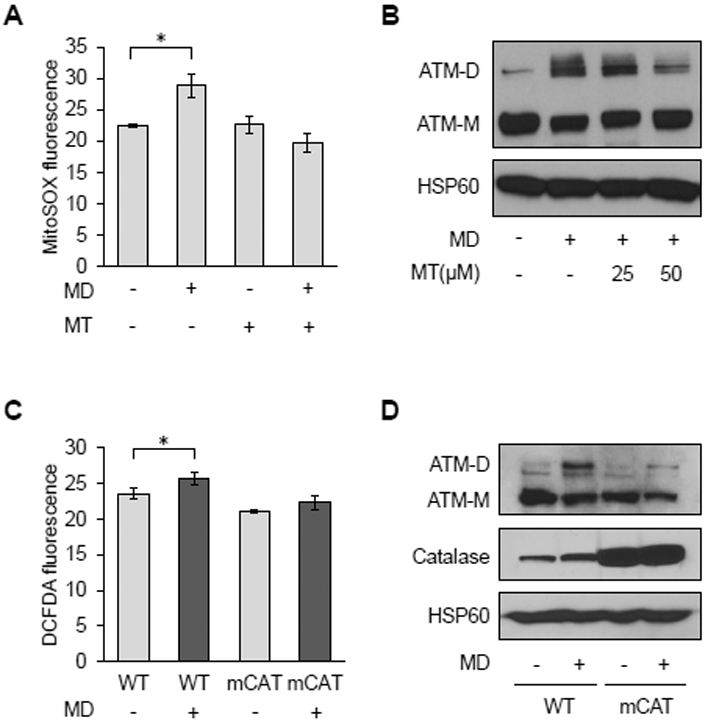
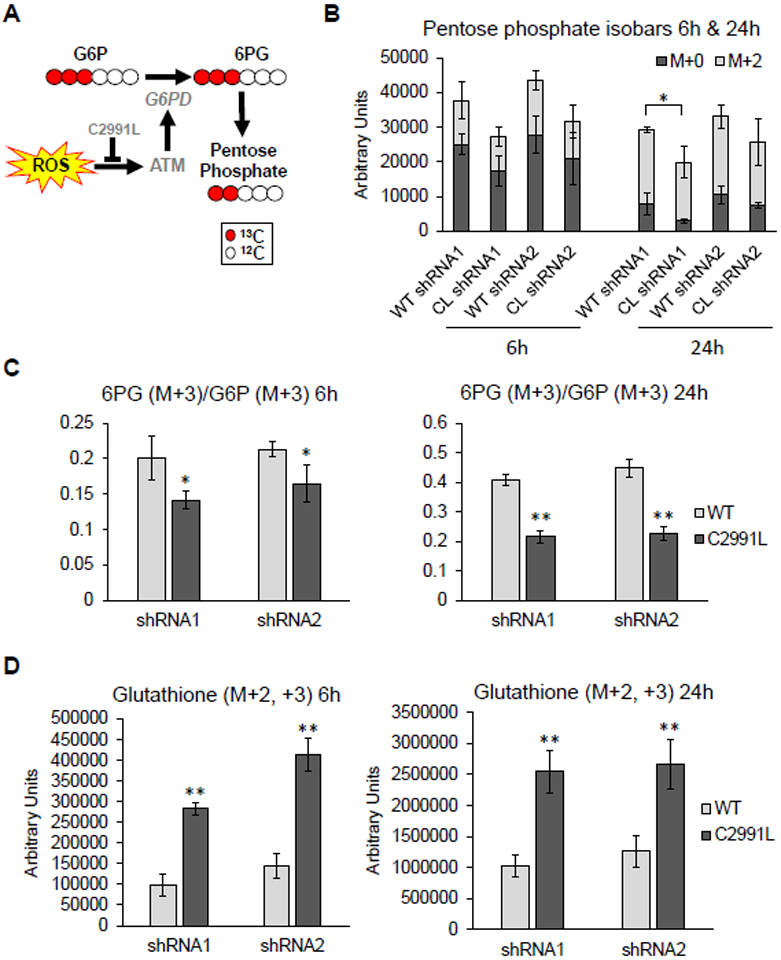
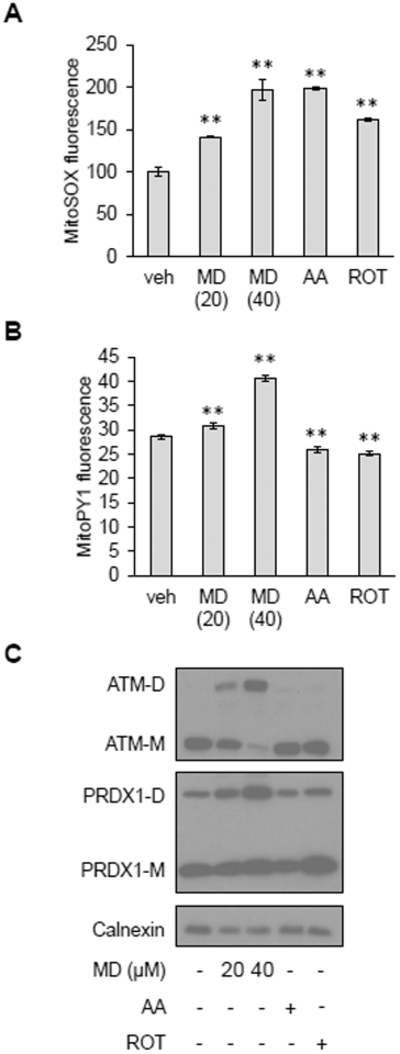
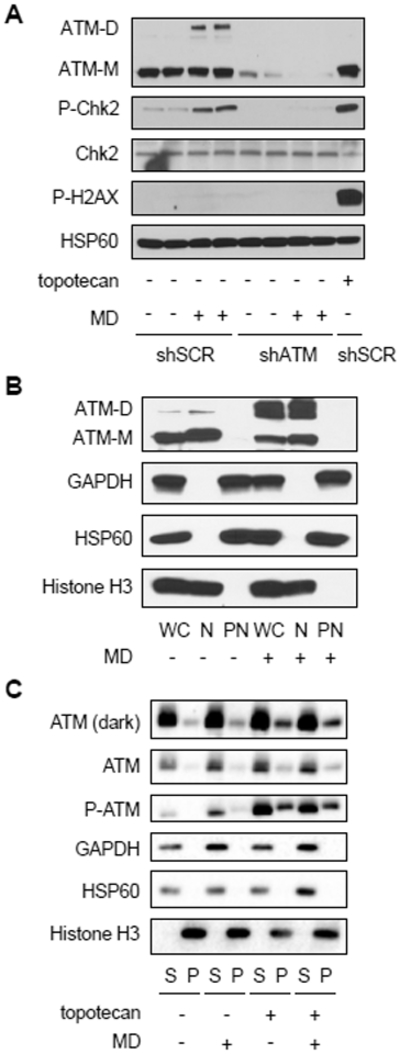
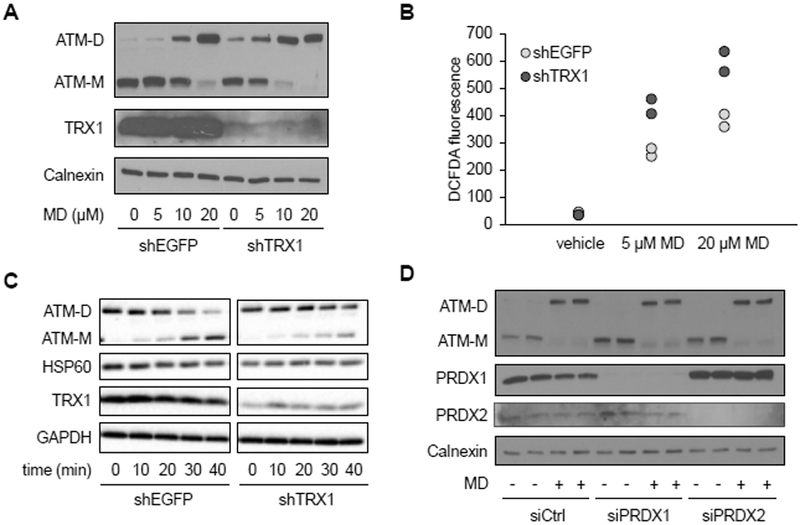
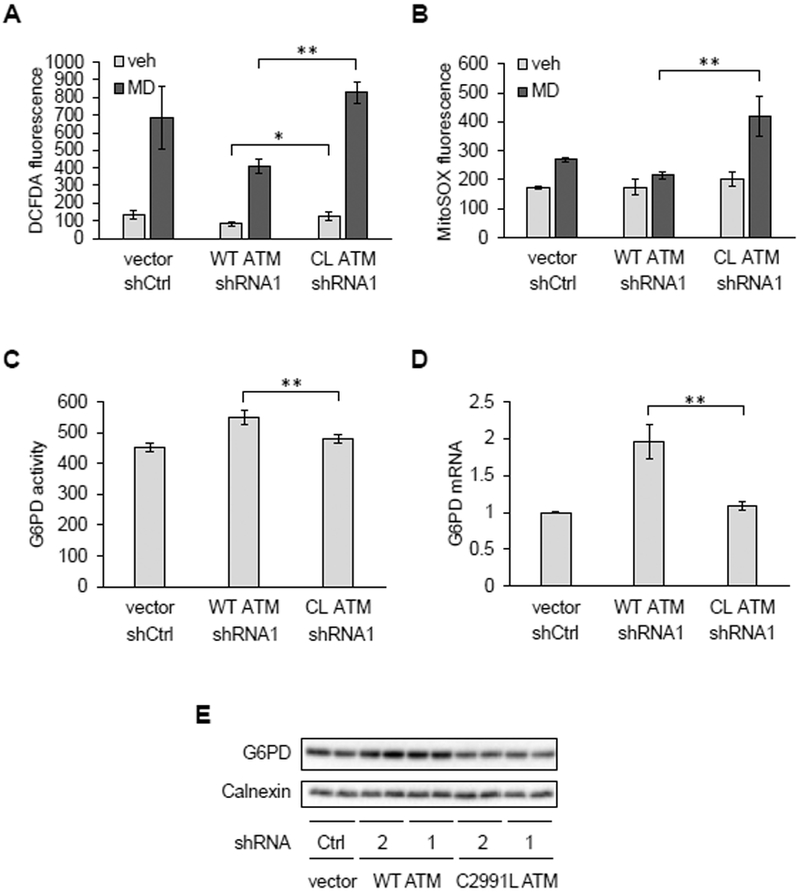
Mitochondria are integral to cellular energy metabolism and ATP production and are involved in regulating many cellular processes. Mitochondria produce reactive oxygen species (ROS), which not only can damage cellular components but also participate in signal transduction. The kinase ATM, which is mutated in the neurodegenerative, autosomal recessive disease ataxia-telangiectasia (A-T), is a key player in the nuclear DNA damage response. However, ATM also performs a redox-sensing function mediated through formation of ROS-dependent disulfide-linked dimers. We found that mitochondria-derived hydrogen peroxide promoted ATM dimerization. In HeLa cells, ATM dimers were localized to the nucleus and inhibited by the redox regulatory protein thioredoxin 1 (TRX1), suggesting the existence of a ROS-mediated, stress-signaling relay from mitochondria to the nucleus. ATM dimer formation did not affect its association with chromatin in the absence or presence of nuclear DNA damage, consistent with the separation of its redox and DNA damage signaling functions. Comparative analysis of U2OS cells expressing either wild-type ATM or the redox sensing-deficient C2991L mutant revealed that one function of ATM redox sensing is to promote glucose flux through the pentose phosphate pathway (PPP) by increasing the abundance and activity of glucose-6-phosphate dehydrogenase (G6PD), thereby increasing cellular antioxidant capacity. The PPP produces the coenzyme NADPH needed for a robust antioxidant response, including the regeneration of TRX1, indicating the existence of a regulatory feedback loop involving ATM and TRX1. We propose that loss of the mitochondrial ROS-sensing function of ATM may cause cellular ROS accumulation and oxidative stress in A-T.
线粒体是细胞能量代谢和 ATP 产生的重要组成部分,参与调节许多细胞过程。线粒体产生活性氧(ROS),ROS 不仅可以破坏细胞成分,还参与信号转导。激酶 ATM 在神经退行性疾病常染色体隐性遗传共济失调-毛细血管扩张症(A-T)中发生突变,是核 DNA 损伤反应的关键参与者。然而,ATM 还通过形成 ROS 依赖的二硫键连接的二聚体来执行氧化还原感应功能。我们发现,线粒体衍生的过氧化氢促进了 ATM 二聚体的形成。在 HeLa 细胞中,ATM 二聚体定位于细胞核,并被氧化还原调节蛋白硫氧还蛋白 1(TRX1)抑制,这表明存在从线粒体到细胞核的 ROS 介导的应激信号转导中继。ATM 二聚体的形成不影响其与染色质的结合,无论是在没有核 DNA 损伤的情况下还是存在核 DNA 损伤的情况下,这与它的氧化还原和 DNA 损伤信号功能的分离一致。对表达野生型 ATM 或氧化还原感应缺陷型 C2991L 突变体的 U2OS 细胞进行比较分析表明,ATM 氧化还原感应的一个功能是通过增加葡萄糖-6-磷酸脱氢酶(G6PD)的丰度和活性来促进葡萄糖通过戊糖磷酸途径(PPP)的通量,从而增加细胞抗氧化能力。PPP 产生了强有力的抗氧化反应所需的辅酶 NADPH,包括 TRX1 的再生,这表明存在涉及 ATM 和 TRX1 的调节反馈回路。我们提出,ATM 失去对线粒体 ROS 的感应功能可能导致 A-T 中细胞 ROS 积累和氧化应激。