Nicklin M J, Kräusslich H G, Toyoda H, Dunn J J, Wimmer E
Proc Natl Acad Sci U S A. 1987 Jun;84(12):4002-6. doi: 10.1073/pnas.84.12.4002.
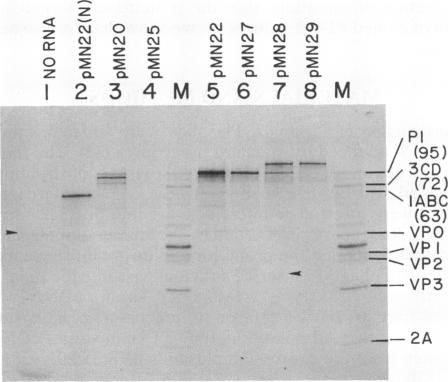
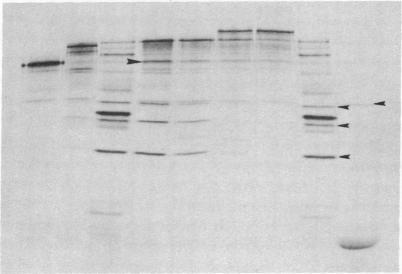
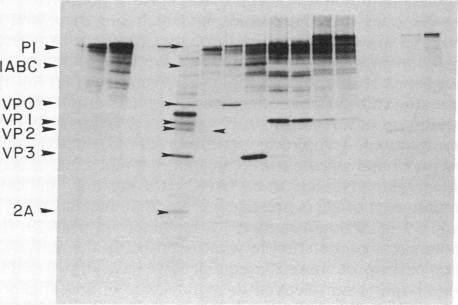
Plasmids have been constructed to generate substrates for the study of proteinases 2A and 3C of poliovirus. They contain the P1 (capsomer precursor) region of the poliovirus genome or P1 and part of P2 (a nonstructural precursor), which can be transcribed and translated in vitro. A transcript containing the entire 5' nontranslated region and the P1 region of the viral RNA gave poor translation in a reticulocyte translation system. Truncation of the 5' nontranslated region to its 3'-most segment gave acceptably good yields of radiolabeled P1. P1 was specifically processed to yield capsomer proteins by enzymes supplied in a postmitochondrial supernatant from poliovirus-infected cells. Thus, proteinase 3C can be supplied exogenously (in trans) and effect processing. This system may be used to provide P1 for the assay of proteinase 3C. Precursors that lacked either the 1A or 1D regions were poor substrates for proteinase 3C--observations that demonstrated a stringent structural requirement in processing by 3C. The translation product of a transcript encoding P1 and part of P2 was rapidly cleaved at the P1-P2 site in the absence of infected-cell extract. A transcript that contained a mutated 2A region gave a stable P1-P2 precursor that could be processed specifically by exogenous proteinase from infected-cell fractions. Processing of P1 appeared to require cleavage of the P1-P2 bond. These results support our previous data that 2A is the second polioviral proteinase and also provides a means of assaying proteinase 2A in vitro.
已构建质粒以生成用于研究脊髓灰质炎病毒蛋白酶2A和3C的底物。它们包含脊髓灰质炎病毒基因组的P1(衣壳蛋白前体)区域或P1和部分P2(一种非结构前体),可在体外进行转录和翻译。在网织红细胞翻译系统中,包含病毒RNA完整5'非翻译区和P1区域的转录本翻译效果不佳。将5'非翻译区截短至其最3'端片段,可获得放射性标记P1的良好产量。P1通过脊髓灰质炎病毒感染细胞的线粒体后上清液中提供的酶进行特异性加工,产生衣壳蛋白。因此,蛋白酶3C可外源提供(反式)并发挥加工作用。该系统可用于提供P1以检测蛋白酶3C。缺少1A或1D区域的前体是蛋白酶3C的不良底物——这些观察结果表明3C加工存在严格的结构要求。在没有感染细胞提取物的情况下,编码P1和部分P2的转录本的翻译产物在P1 - P2位点迅速裂解。包含突变2A区域的转录本产生稳定的P1 - P2前体,可被感染细胞组分中的外源蛋白酶特异性加工。P1的加工似乎需要P1 - P2键的裂解。这些结果支持了我们之前的数据,即2A是第二种脊髓灰质炎病毒蛋白酶,也提供了一种体外检测蛋白酶2A的方法。