Janelia Research Campus, Howard Hughes Medical Institute, Ashburn, United States.
Elife. 2018 Oct 30;7:e37701. doi: 10.7554/eLife.37701.
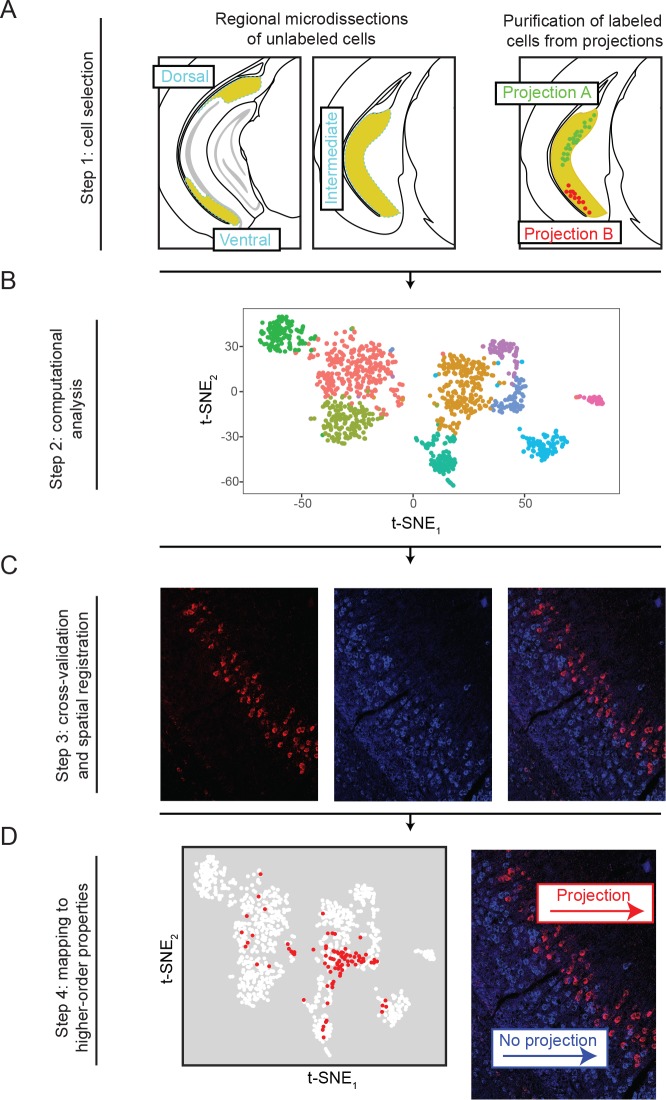
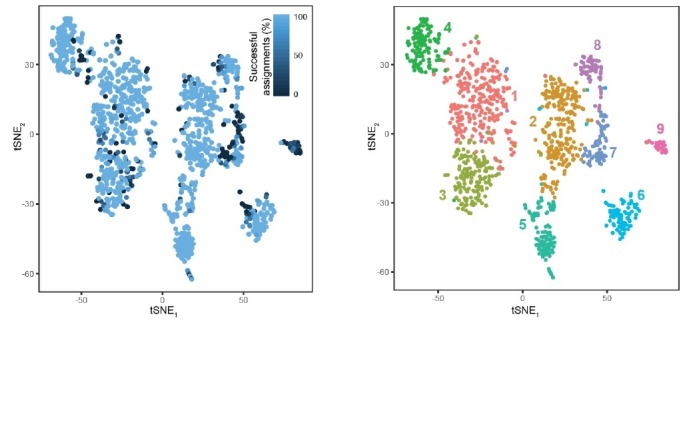
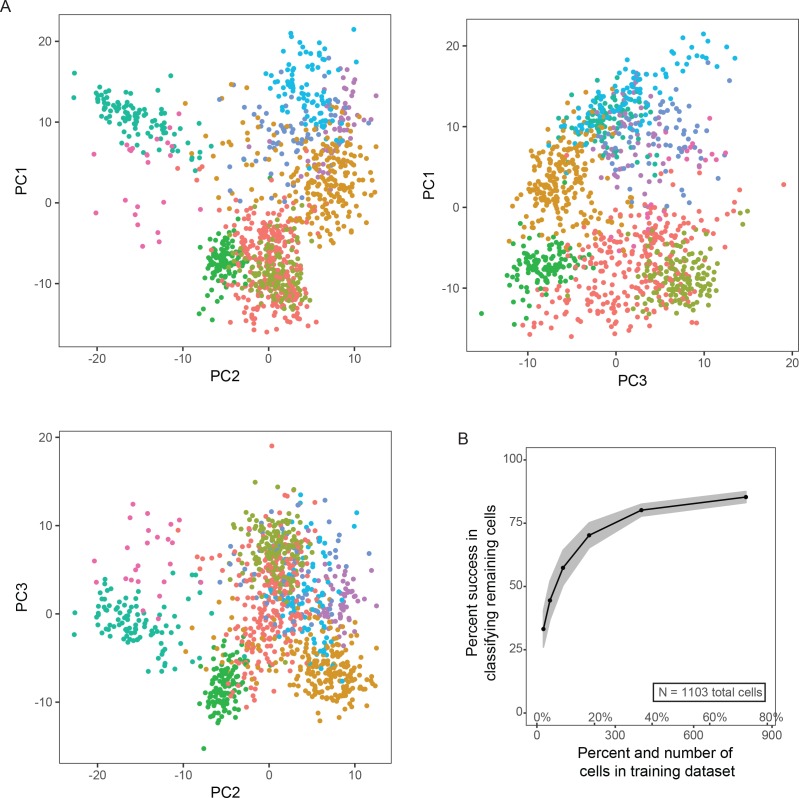
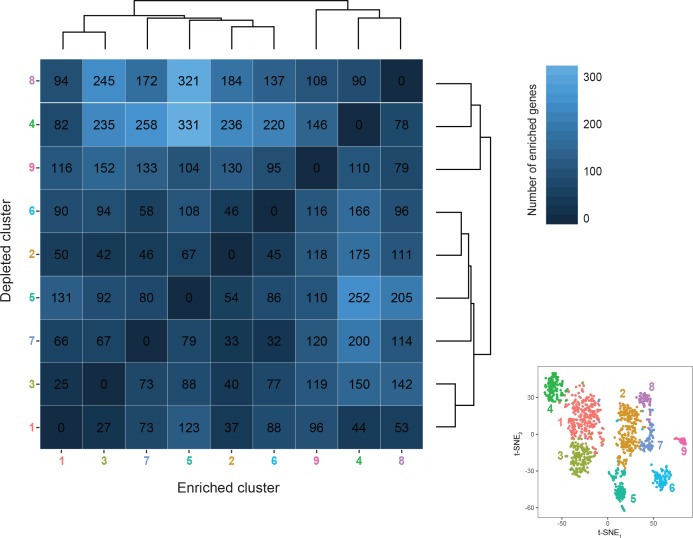
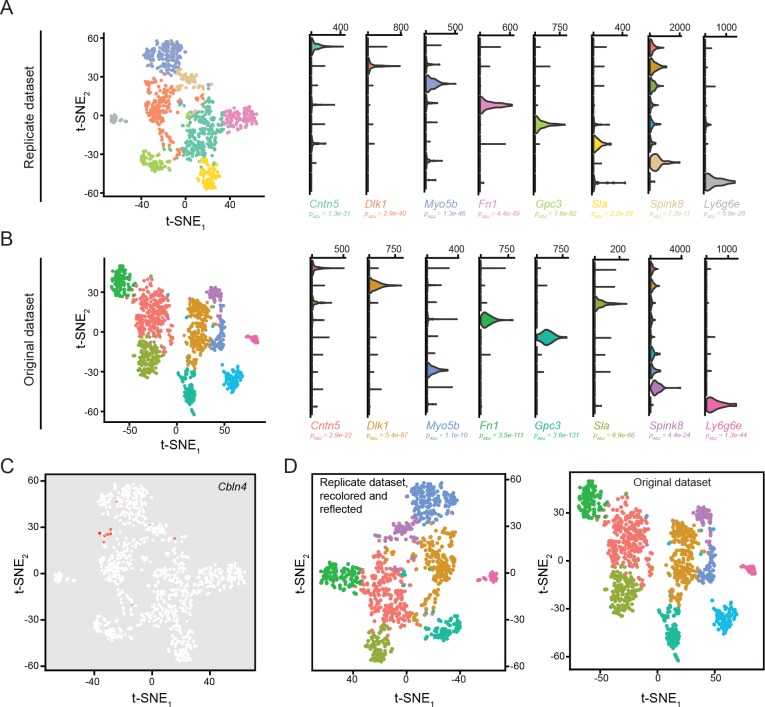
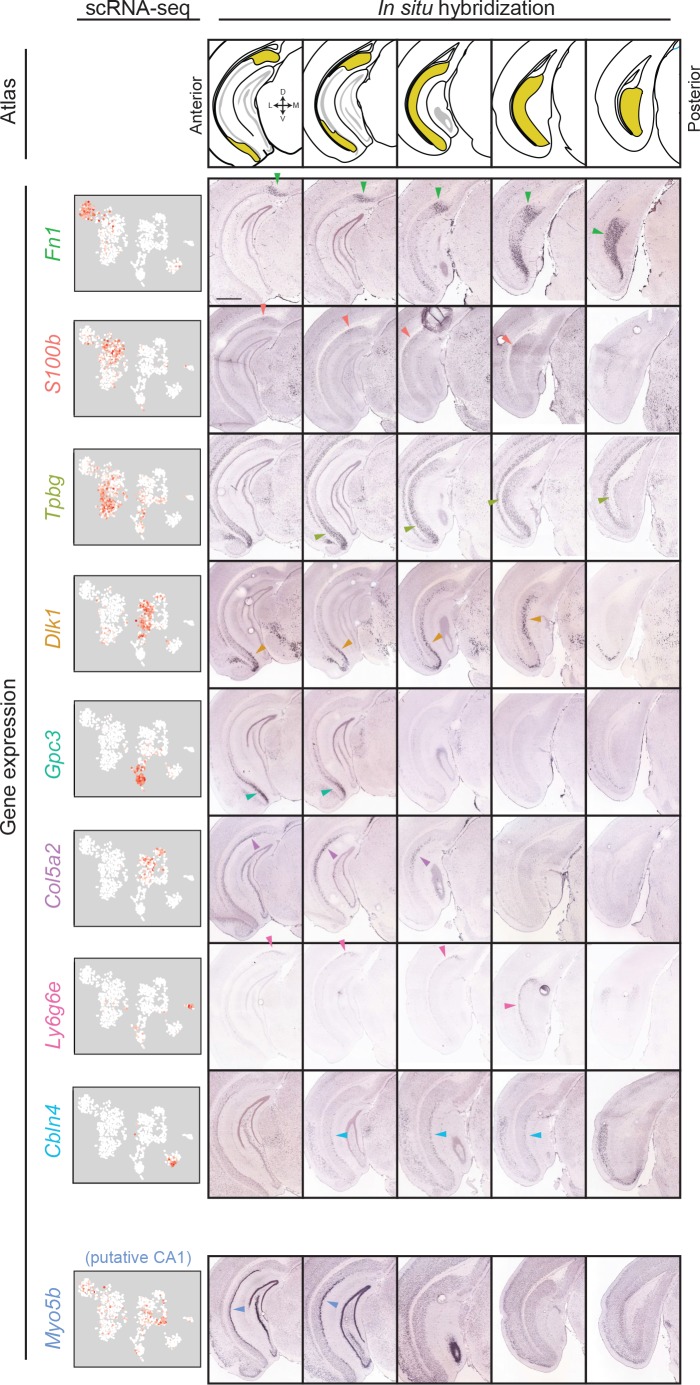
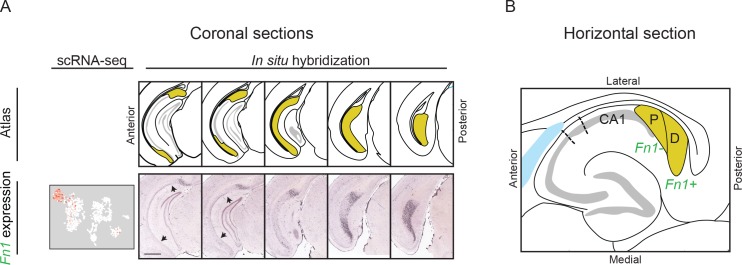
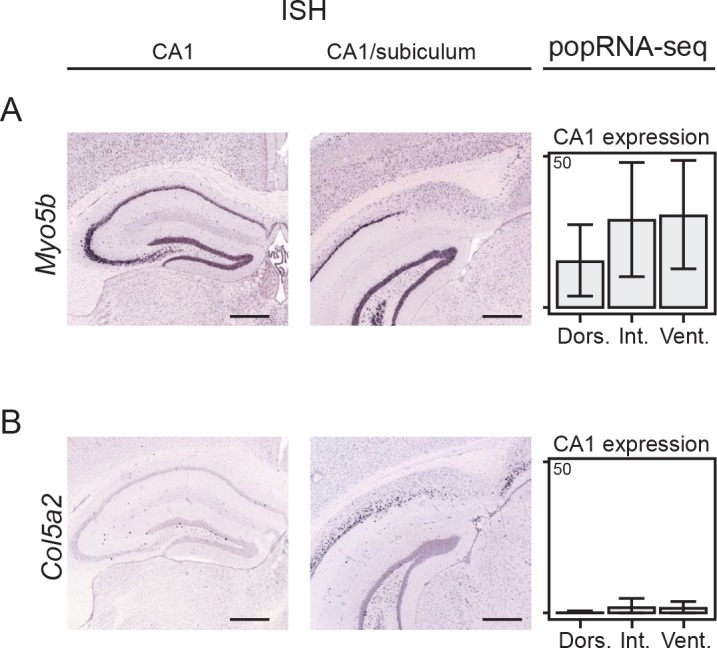
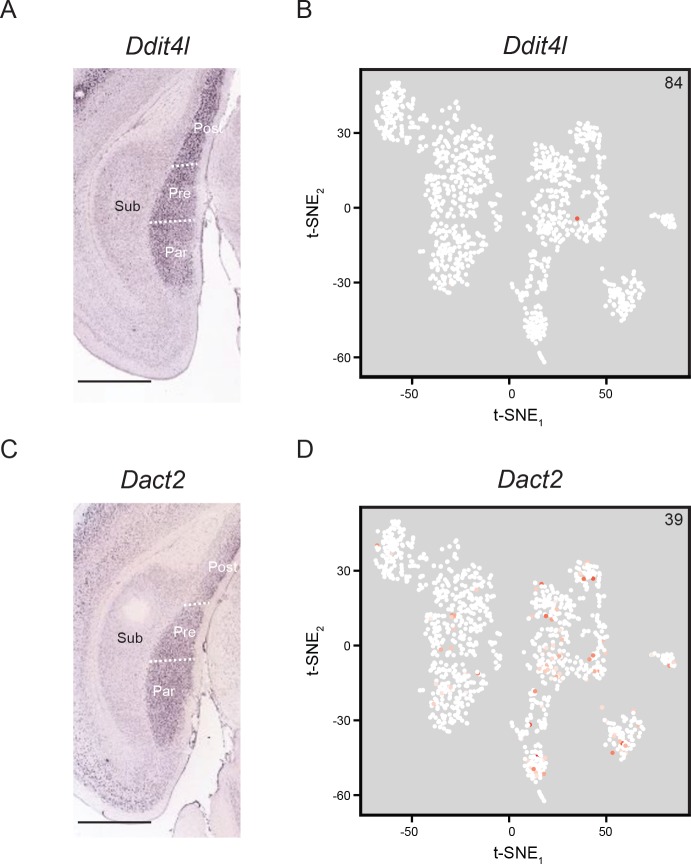
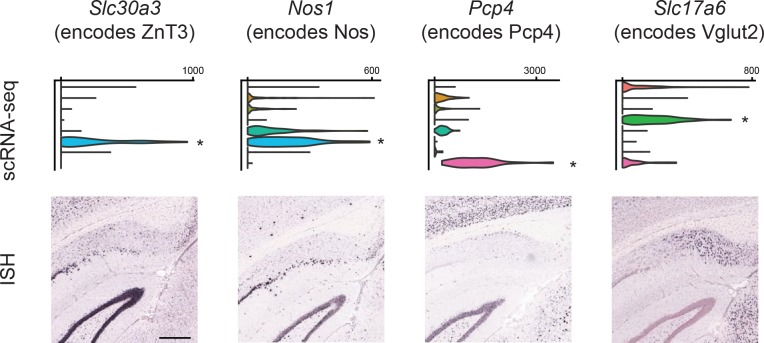
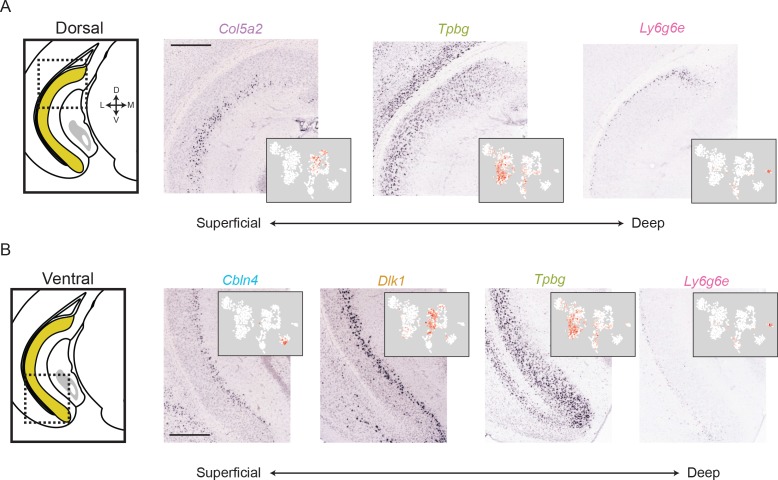
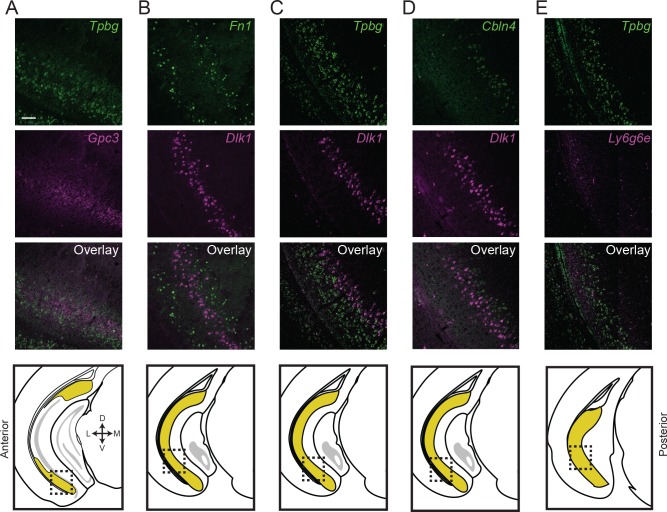
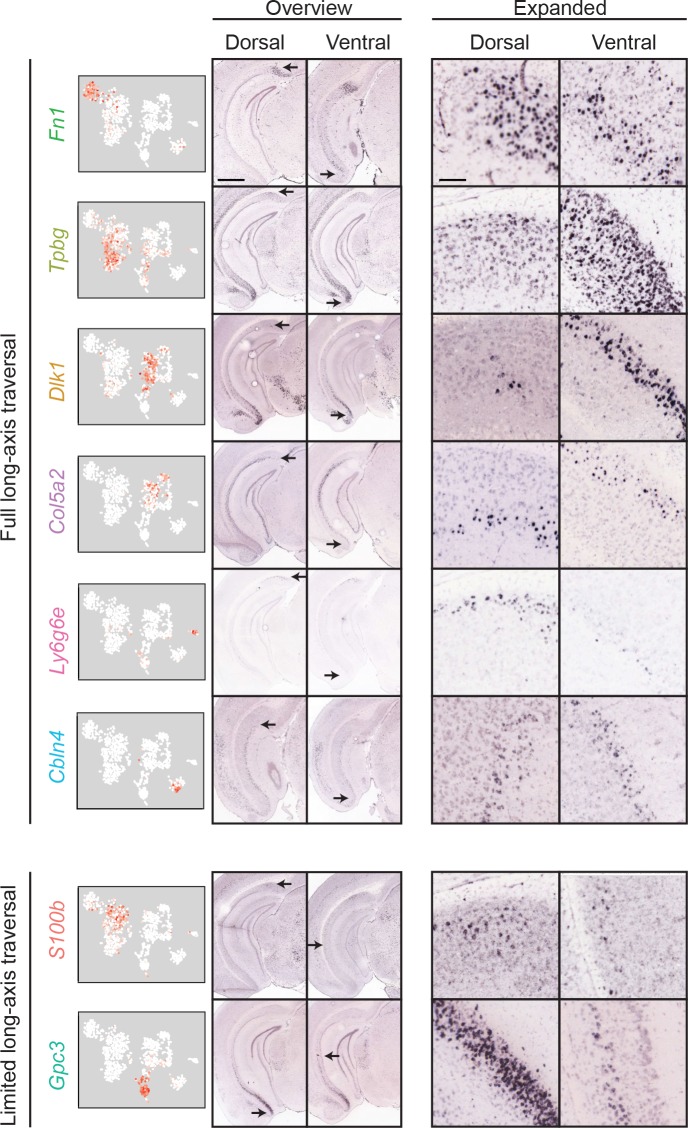
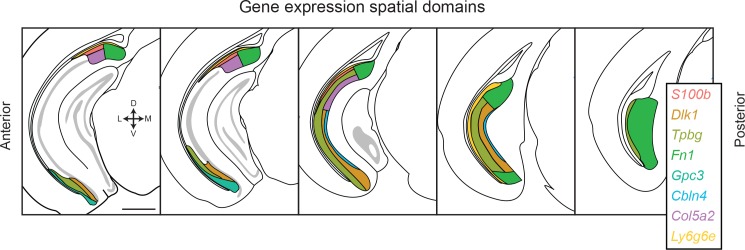
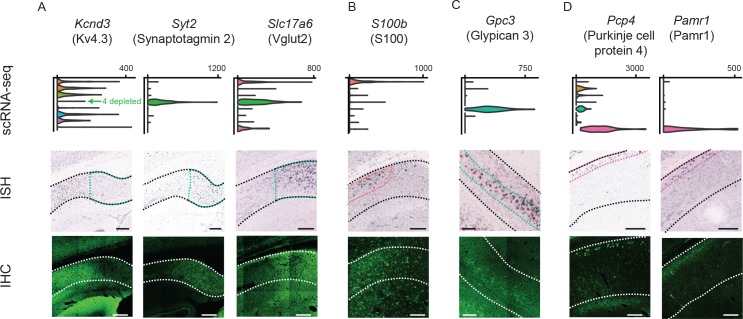
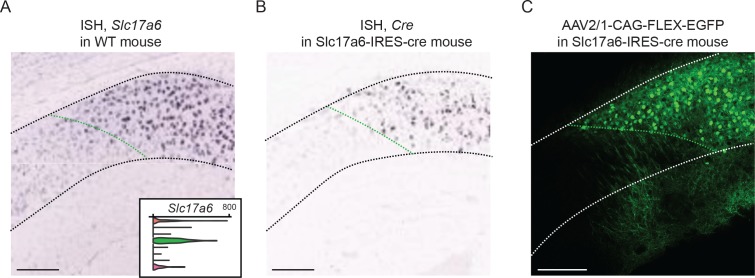
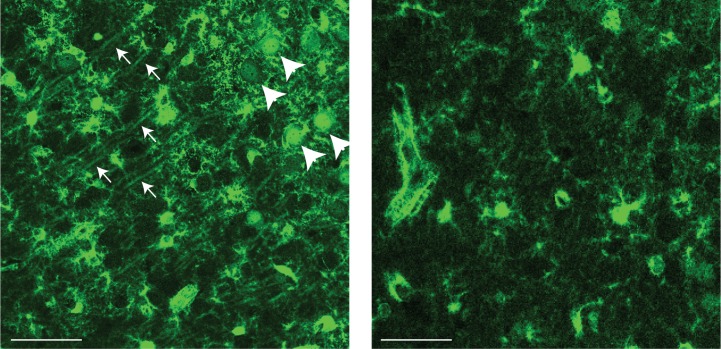
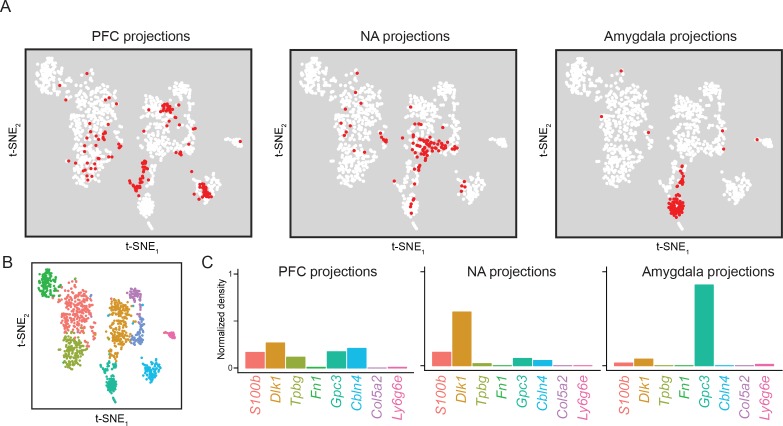
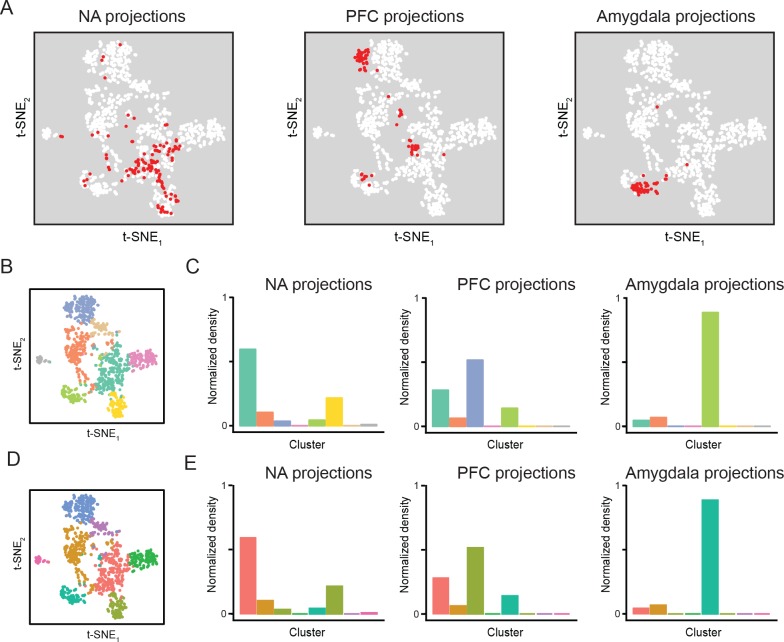
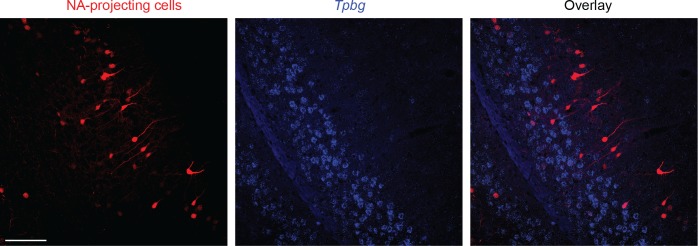
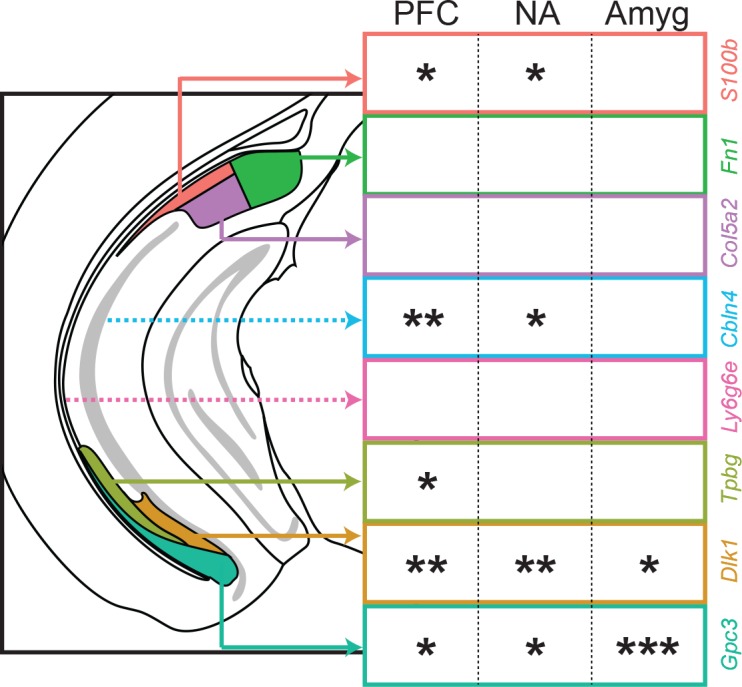
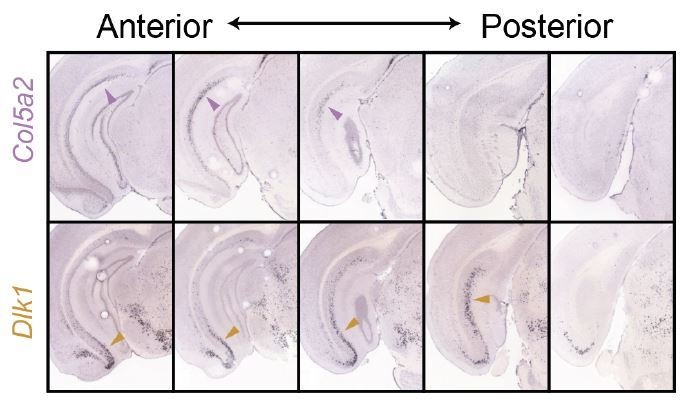
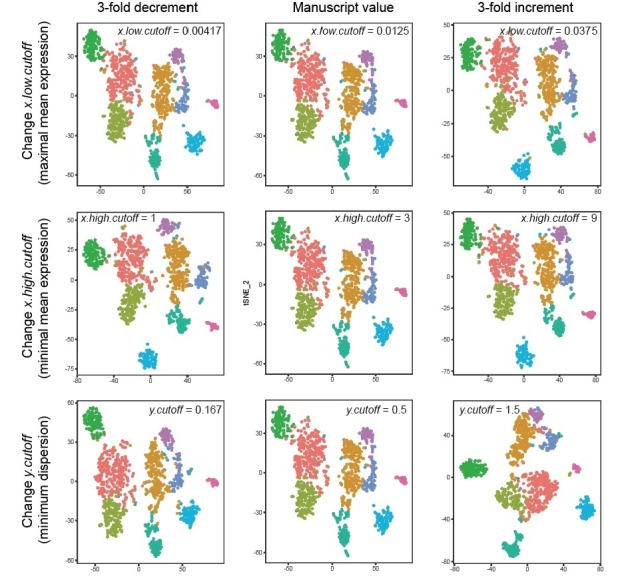
In the hippocampus, the classical pyramidal cell type of the subiculum acts as a primary output, conveying hippocampal signals to a diverse suite of downstream regions. Accumulating evidence suggests that the subiculum pyramidal cell population may actually be comprised of discrete subclasses. Here, we investigated the extent and organizational principles governing pyramidal cell heterogeneity throughout the mouse subiculum. Using single-cell RNA-seq, we find that the subiculum pyramidal cell population can be deconstructed into eight separable subclasses. These subclasses were mapped onto abutting spatial domains, ultimately producing a complex laminar and columnar organization with heterogeneity across classical dorsal-ventral, proximal-distal, and superficial-deep axes. We further show that these transcriptomically defined subclasses correspond to differential protein products and can be associated with specific projection targets. This work deconstructs the complex landscape of subiculum pyramidal cells into spatially segregated subclasses that may be observed, controlled, and interpreted in future experiments.
在海马体中,作为主要输出的经典下托锥体神经元类型将海马体信号传递到各种下游区域。越来越多的证据表明,下托锥体神经元群体实际上可能由离散的亚类组成。在这里,我们研究了控制整个小鼠下托中锥体神经元异质性的程度和组织原则。通过单细胞 RNA 测序,我们发现下托锥体神经元群体可以被分解为八个可分离的亚类。这些亚类被映射到相邻的空间域上,最终产生了一个复杂的层状和柱状组织,具有跨经典背腹、近远和浅深轴的异质性。我们进一步表明,这些转录定义的亚类对应于不同的蛋白质产物,并可以与特定的投射靶标相关联。这项工作将下托锥体神经元的复杂景观分解为空间上分离的亚类,这些亚类可能在未来的实验中被观察、控制和解释。