Laboratoire Physico Chimie Curie, Institut Curie, PSL Research University, Sorbonne Université, CNRS, 75005, Paris, France.
PASTEUR, Département de chimie, École normale supérieure, CNRS UMR 8640, PSL Research University, Sorbonne Université, 75005, Paris, France.
Nat Commun. 2018 Nov 16;9(1):4816. doi: 10.1038/s41467-018-07286-8.
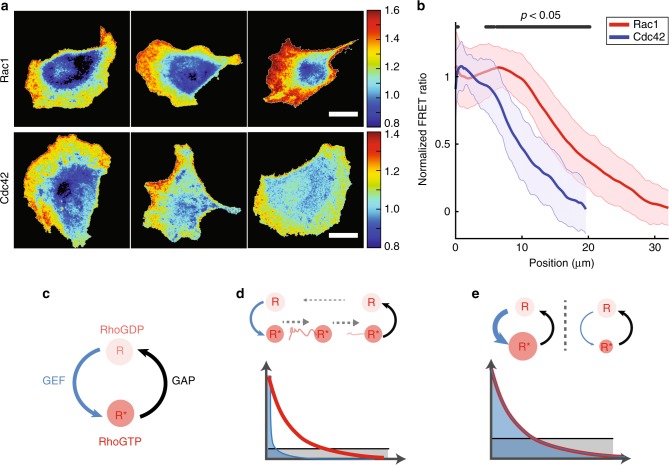
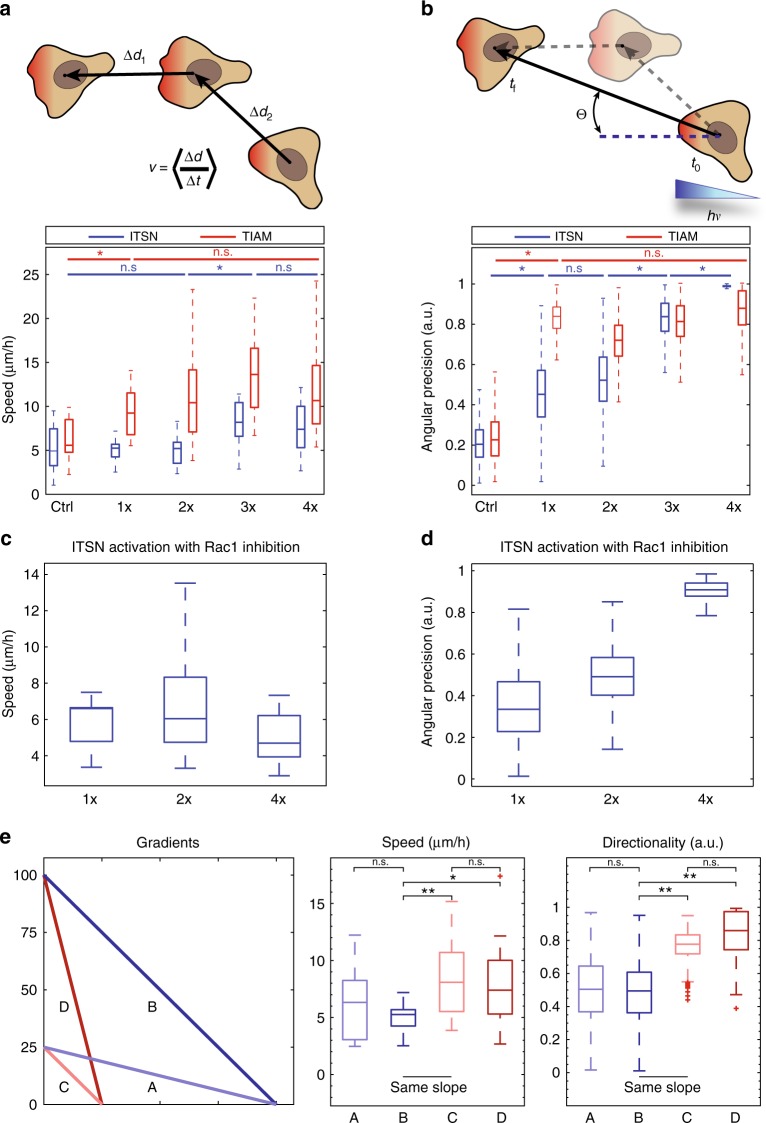
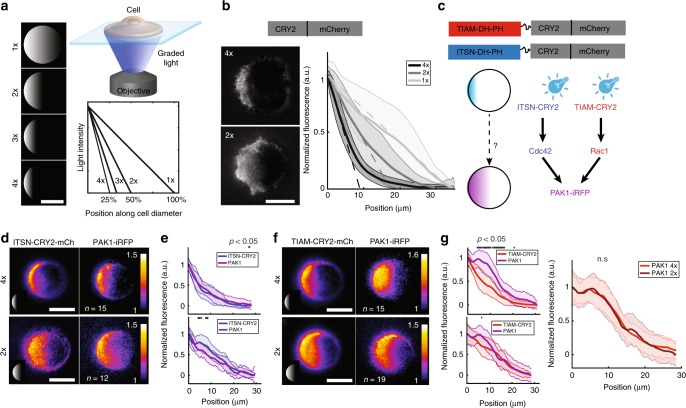
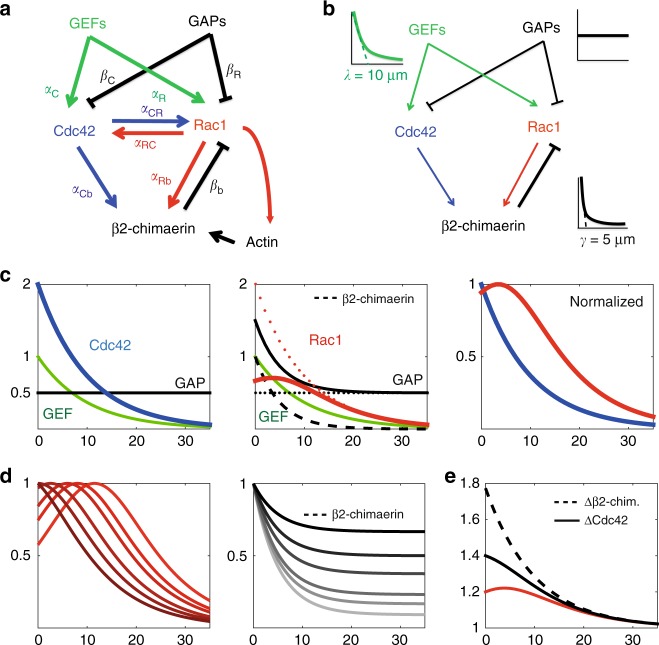
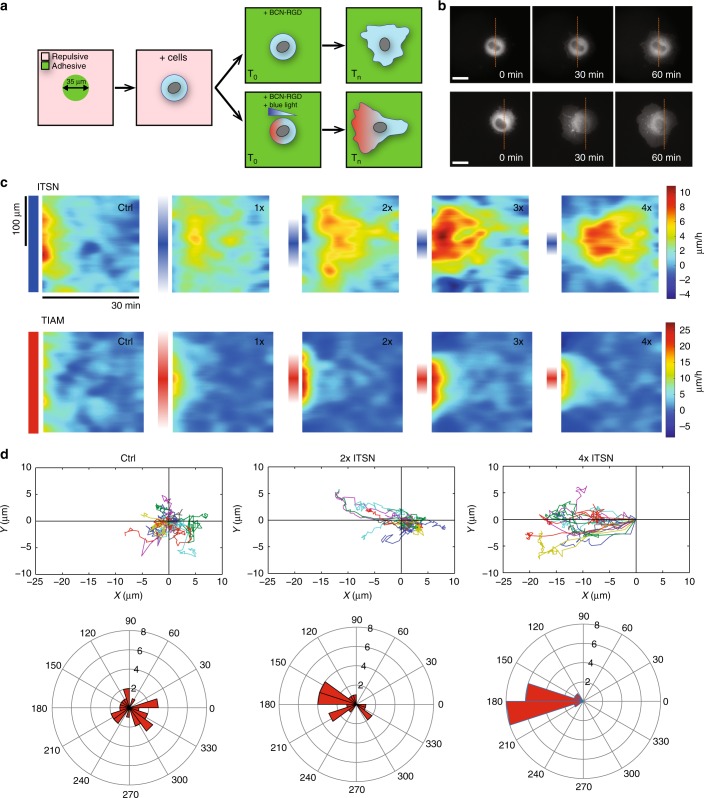
During cell migration, Rho GTPases spontaneously form spatial gradients that define the front and back of cells. At the front, active Cdc42 forms a steep gradient whereas active Rac1 forms a more extended pattern peaking a few microns away. What are the mechanisms shaping these gradients, and what is the functional role of the shape of these gradients? Here we report, using a combination of optogenetics and micropatterning, that Cdc42 and Rac1 gradients are set by spatial patterns of activators and deactivators and not directly by transport mechanisms. Cdc42 simply follows the distribution of Guanine nucleotide Exchange Factors, whereas Rac1 shaping requires the activity of a GTPase-Activating Protein, β2-chimaerin, which is sharply localized at the tip of the cell through feedbacks from Cdc42 and Rac1. Functionally, the spatial extent of Rho GTPases gradients governs cell migration, a sharp Cdc42 gradient maximizes directionality while an extended Rac1 gradient controls the speed.
在细胞迁移过程中,Rho GTPases 会自发形成空间梯度,从而定义细胞的前后端。在前部,活性 Cdc42 形成陡峭的梯度,而活性 Rac1 则形成更扩展的模式,在几微米远处达到峰值。形成这些梯度的机制是什么,以及这些梯度的形状有什么功能作用?在这里,我们使用光遗传学和微图案化相结合的方法报告说,Cdc42 和 Rac1 梯度是由激活剂和失活剂的空间模式设定的,而不是直接由运输机制设定的。Cdc42 只是跟随鸟嘌呤核苷酸交换因子的分布,而 Rac1 的形成需要 GTPase 激活蛋白β2-奇美拉的活性,β2-奇美拉通过来自 Cdc42 和 Rac1 的反馈在细胞的尖端被精确地本地化。从功能上讲,Rho GTPases 梯度的空间范围控制着细胞迁移,陡峭的 Cdc42 梯度最大限度地提高了方向性,而扩展的 Rac1 梯度则控制了速度。