ISEM, Université de Montpellier, CNRS, IRD, EPHE, Montpellier, France.
Mol Biol Evol. 2019 Mar 1;36(3):458-471. doi: 10.1093/molbev/msy243.
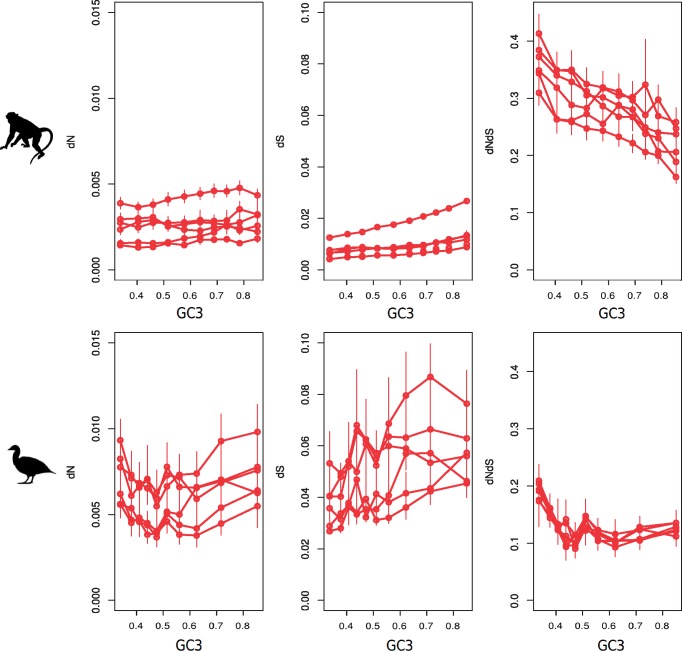
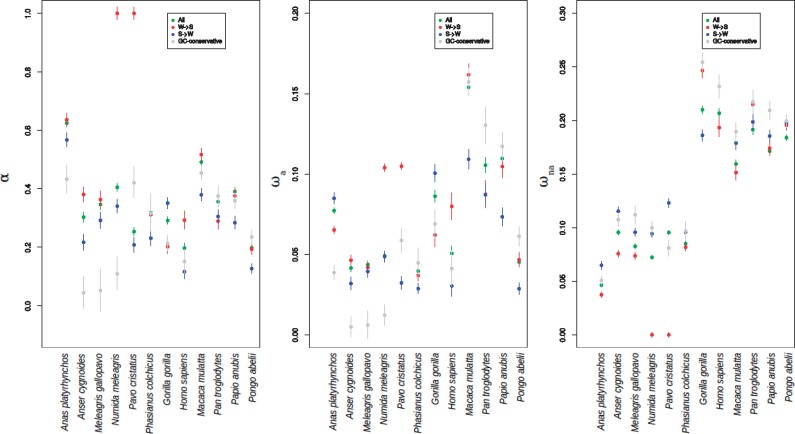
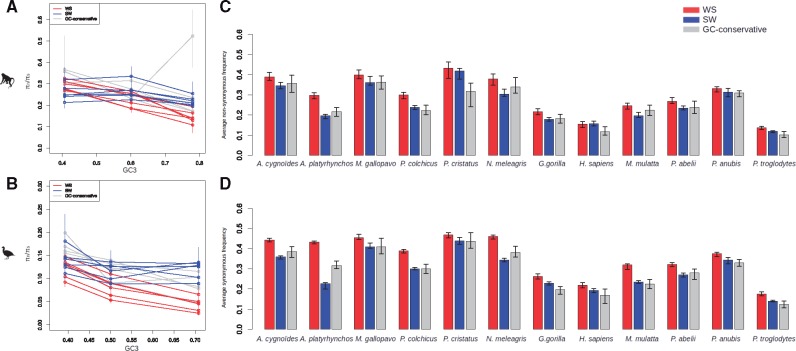
Recombination is expected to affect functional sequence evolution in several ways. On the one hand, recombination is thought to improve the efficiency of multilocus selection by dissipating linkage disequilibrium. On the other hand, natural selection can be counteracted by recombination-associated transmission distorters such as GC-biased gene conversion (gBGC), which tends to promote G and C alleles irrespective of their fitness effect in high-recombining regions. It has been suggested that gBGC might impact coding sequence evolution in vertebrates, and particularly the ratio of nonsynonymous to synonymous substitution rates (dN/dS). However, distinctive gBGC patterns have been reported in mammals and birds, maybe reflecting the documented contrasts in evolutionary dynamics of recombination rate between these two taxa. Here, we explore how recombination and gBGC affect coding sequence evolution in mammals and birds by analyzing proteome-wide data in six species of Galloanserae (fowls) and six species of catarrhine primates. We estimated the dN/dS ratio and rates of adaptive and nonadaptive evolution in bins of genes of increasing recombination rate, separately analyzing AT → GC, GC → AT, and G ↔ C/A ↔ T mutations. We show that in both taxa, recombination and gBGC entail a decrease in dN/dS. Our analysis indicates that recombination enhances the efficiency of purifying selection by lowering Hill-Robertson effects, whereas gBGC leads to an overestimation of the adaptive rate of AT → GC mutations. Finally, we report a mutagenic effect of recombination, which is independent of gBGC.
重组有望通过消除连锁不平衡来影响功能序列进化。一方面,重组被认为可以通过耗散连锁不平衡来提高多点选择的效率。另一方面,自然选择可能会被与重组相关的传输干扰物所抵消,如 GC 偏向性基因转换(gBGC),它倾向于促进 G 和 C 等位基因,而不论其在高重组区域的适应度效应如何。有人认为 gBGC 可能会影响脊椎动物的编码序列进化,特别是非同义替换率与同义替换率(dN/dS)的比值。然而,在哺乳动物和鸟类中已经报道了独特的 gBGC 模式,这可能反映了这两个类群之间重组率进化动态的已有对比。在这里,我们通过分析六种家禽类(禽类)和六种食肉类灵长类动物的全蛋白质组数据,探索了重组和 gBGC 如何影响哺乳动物和鸟类的编码序列进化。我们估计了在基因重组率增加的基因箱中 dN/dS 比值和适应进化与非适应进化的速率,分别分析了 AT→GC、GC→AT 和 G↔C/A↔T 突变。我们表明,在这两个类群中,重组和 gBGC 都导致 dN/dS 的降低。我们的分析表明,重组通过降低 Hill-Robertson 效应来提高纯化选择的效率,而 gBGC 导致对 AT→GC 突变的适应率的高估。最后,我们报告了重组的诱变效应,它独立于 gBGC。