Institute of Biotechnology, HiLIFE/Organismal and Evolutionary Biology Research Programme, Faculty of Biological and Environmental Sciences, Viikki Plant Science Centre, University of Helsinki, Helsinki, Finland.
Graduate School of Science and Technology, Nara Institute of Science and Technology, Nara, Japan.
Nature. 2019 Jan;565(7740):490-494. doi: 10.1038/s41586-018-0839-y. Epub 2019 Jan 9.
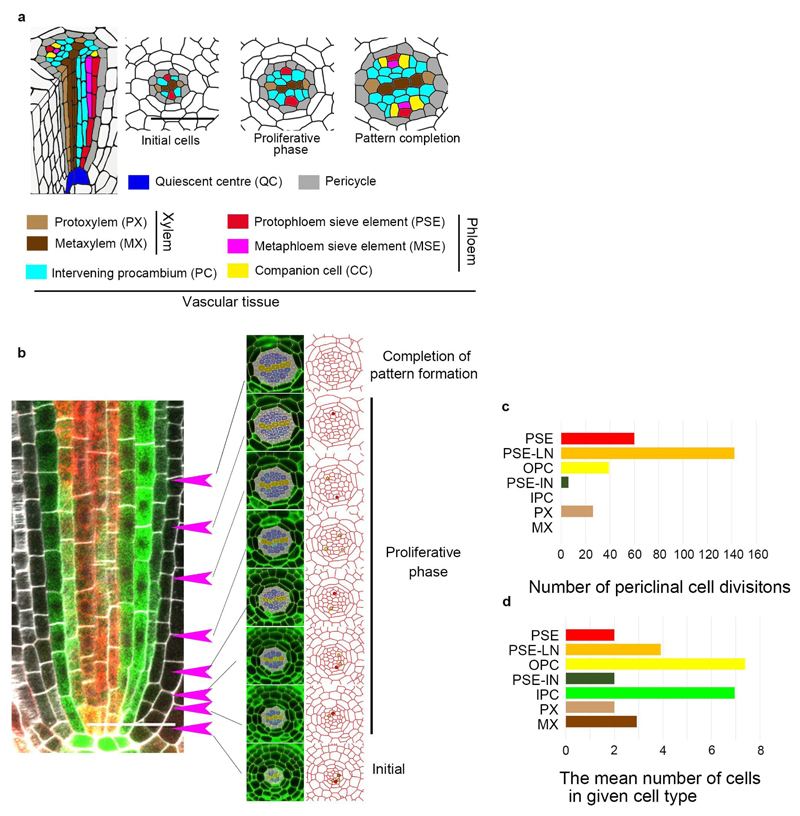
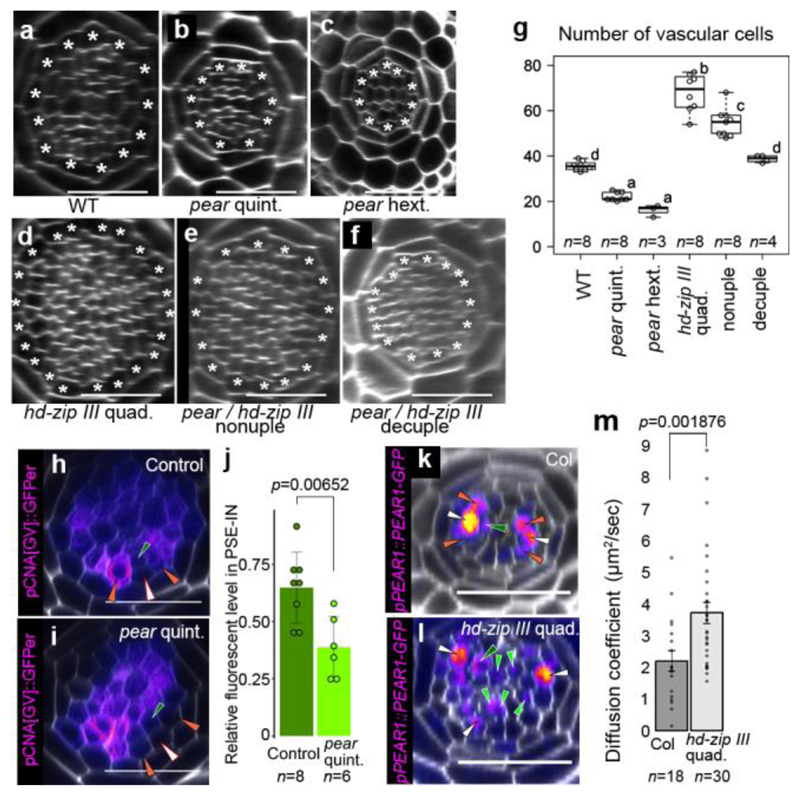
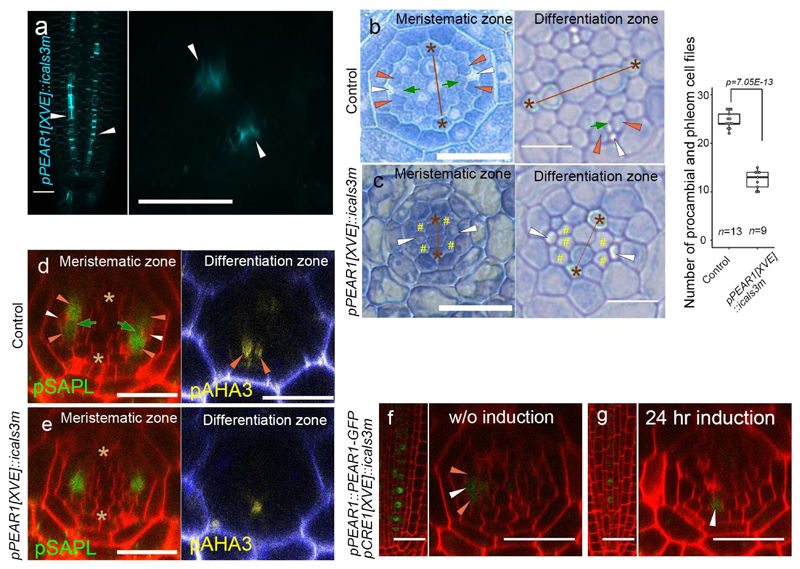
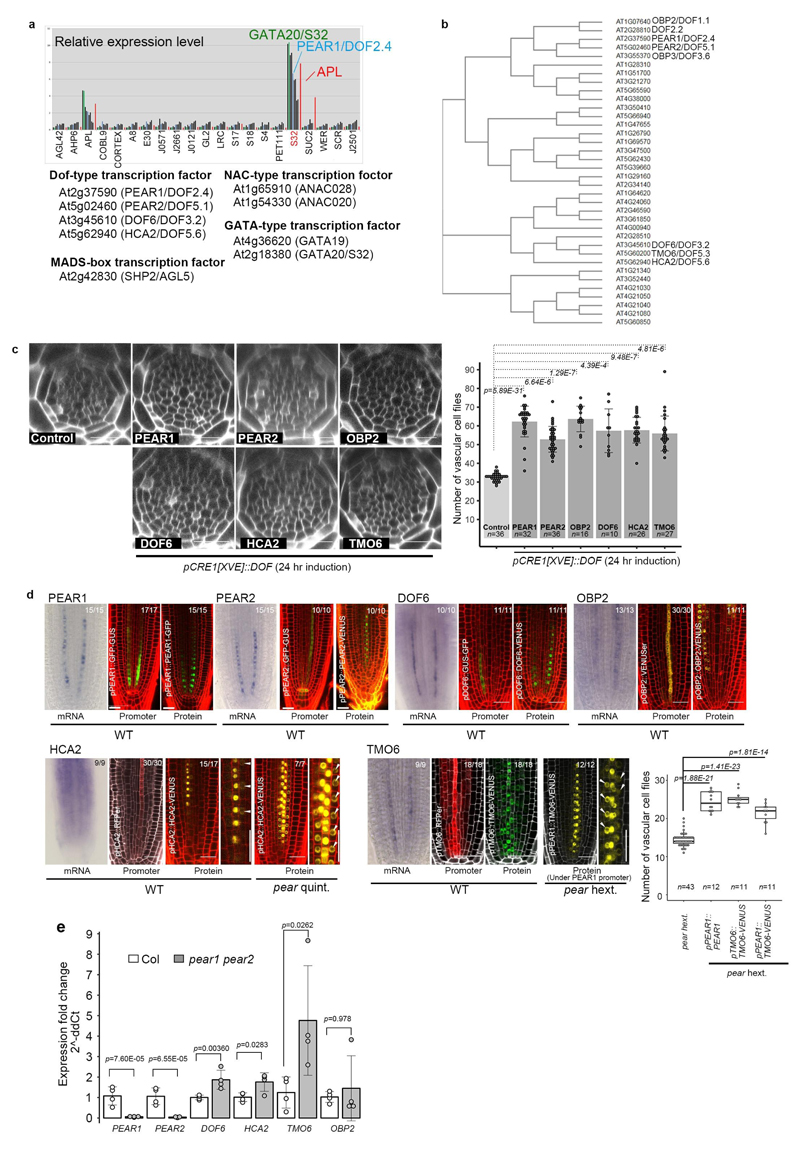
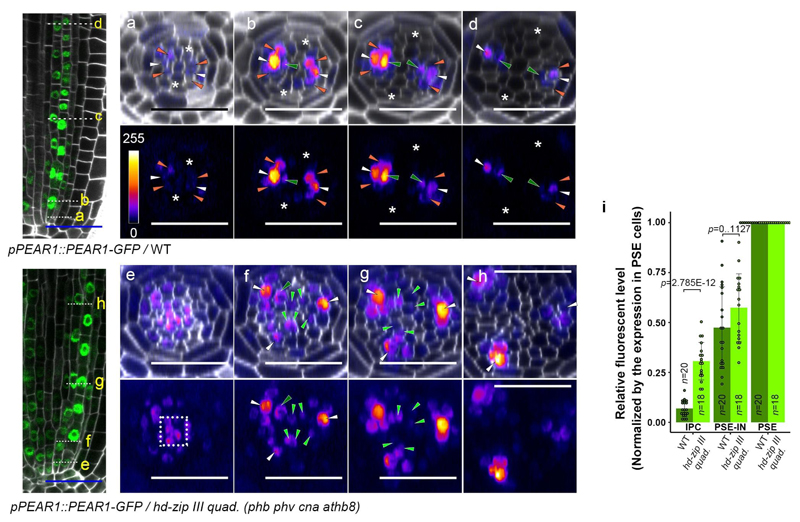
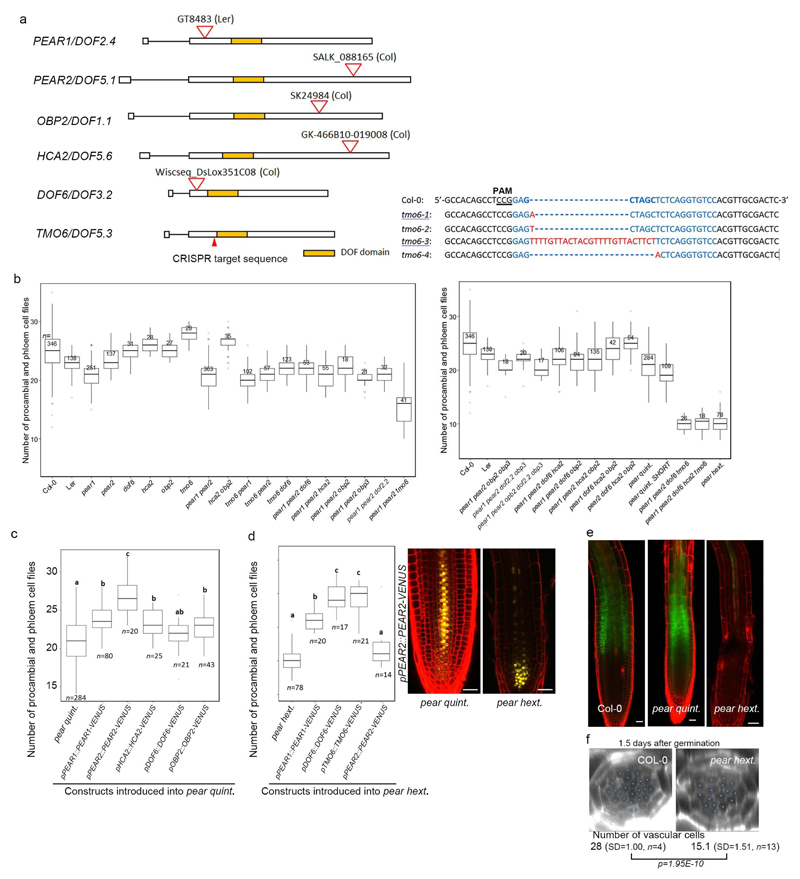
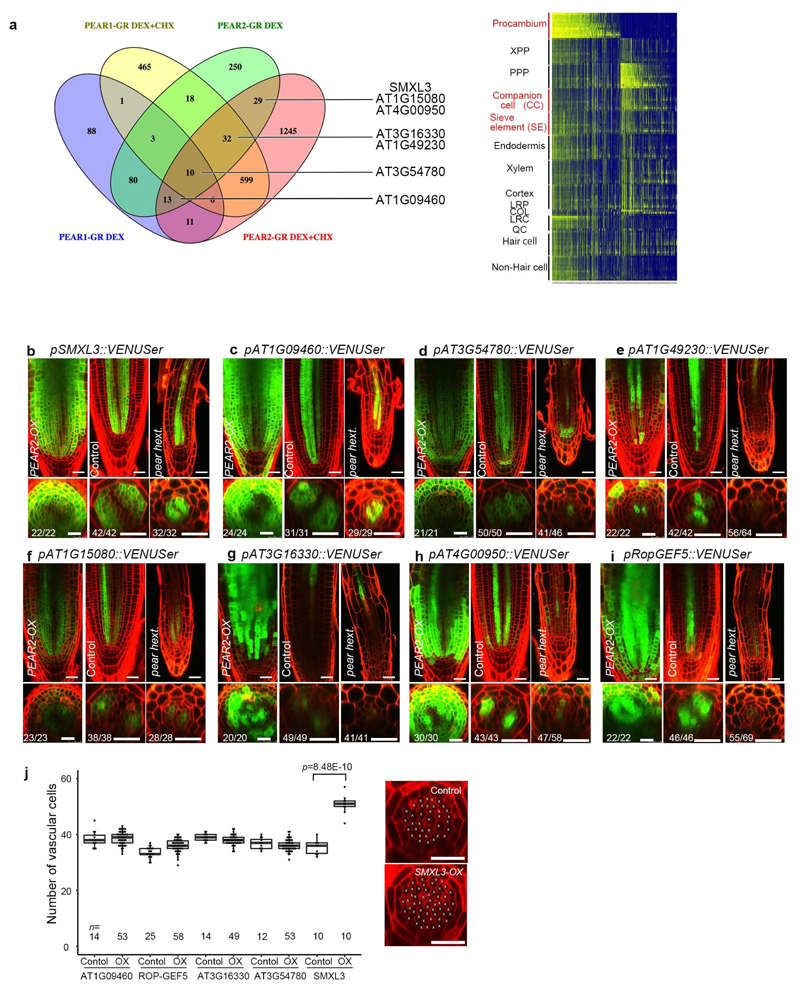
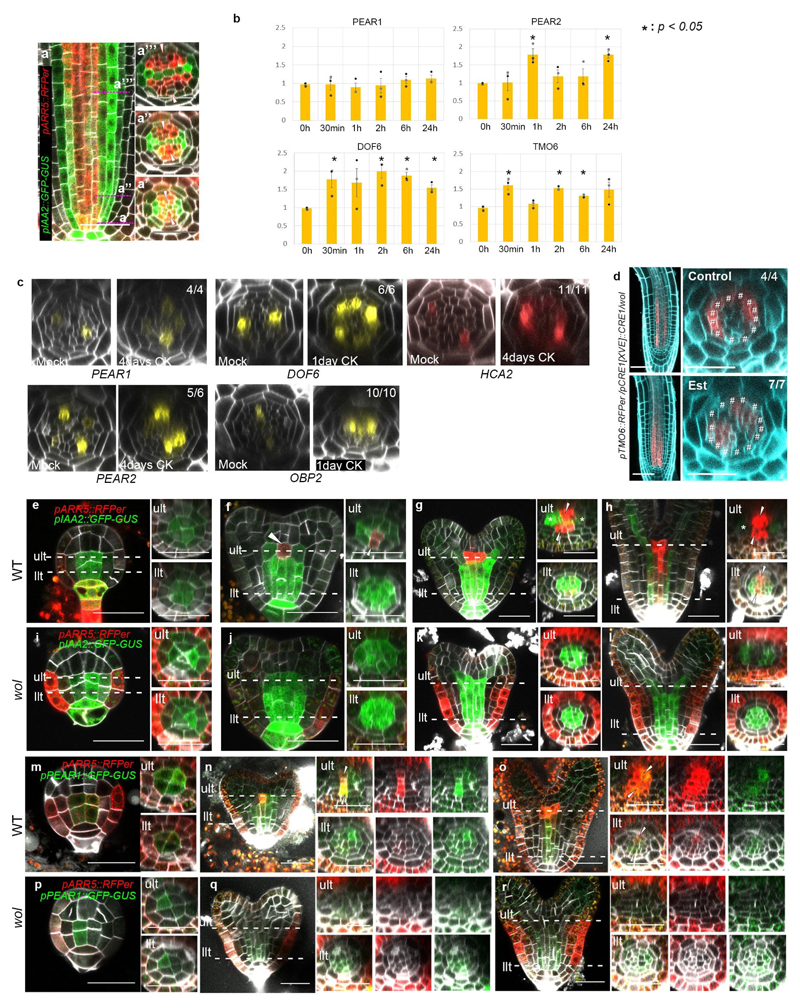
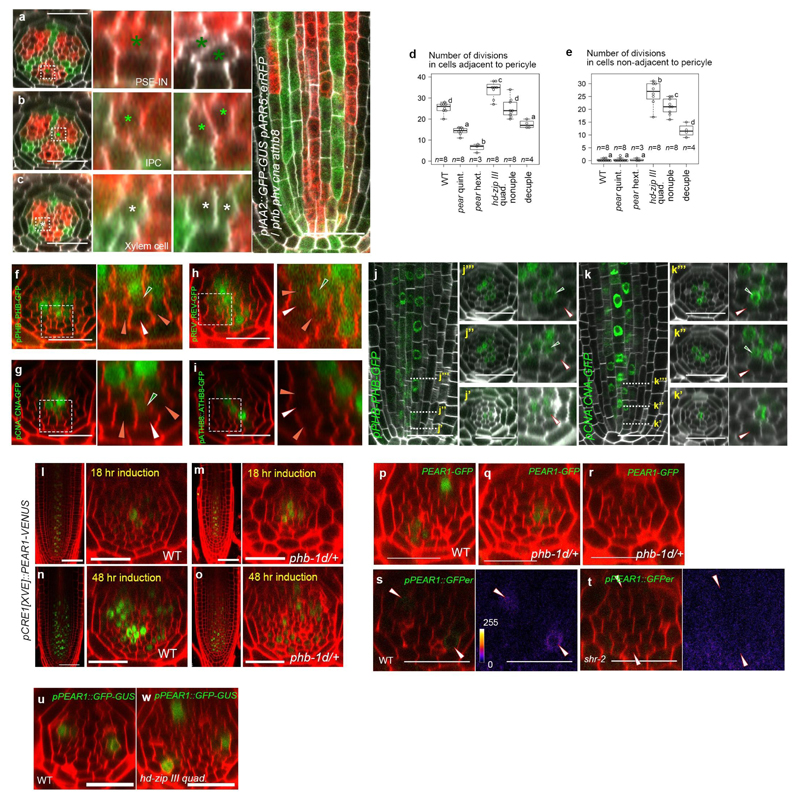
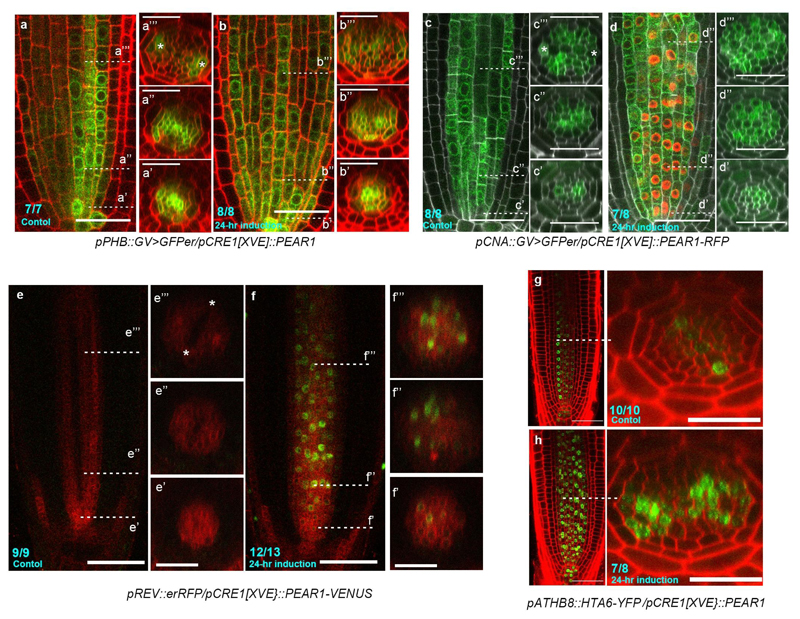
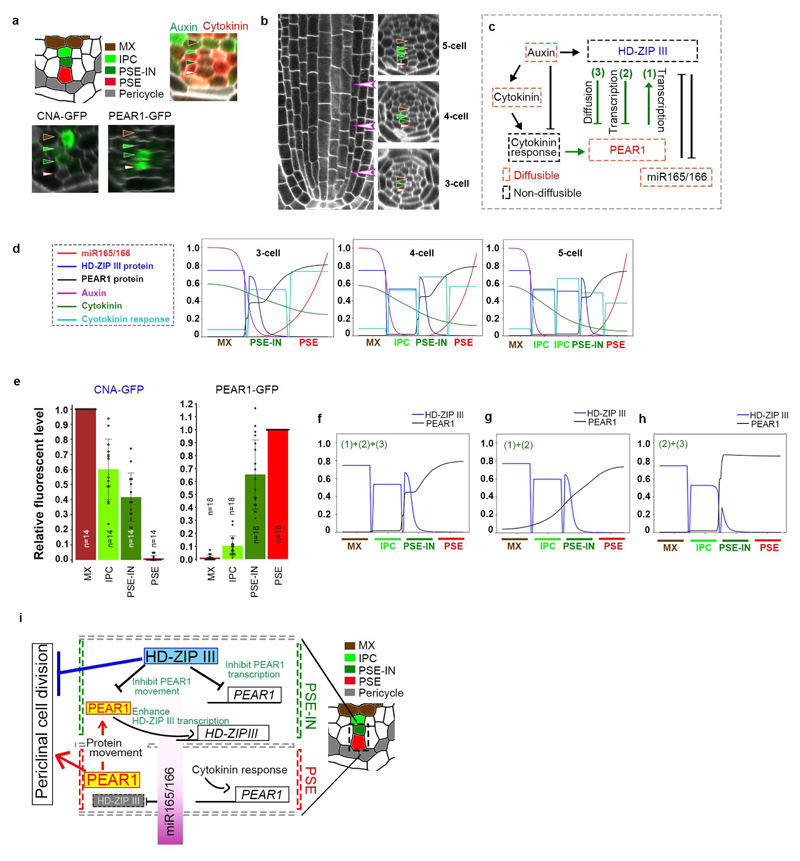
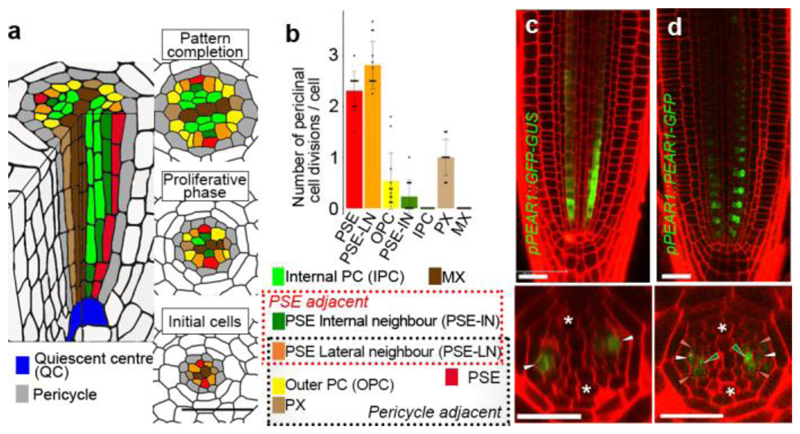
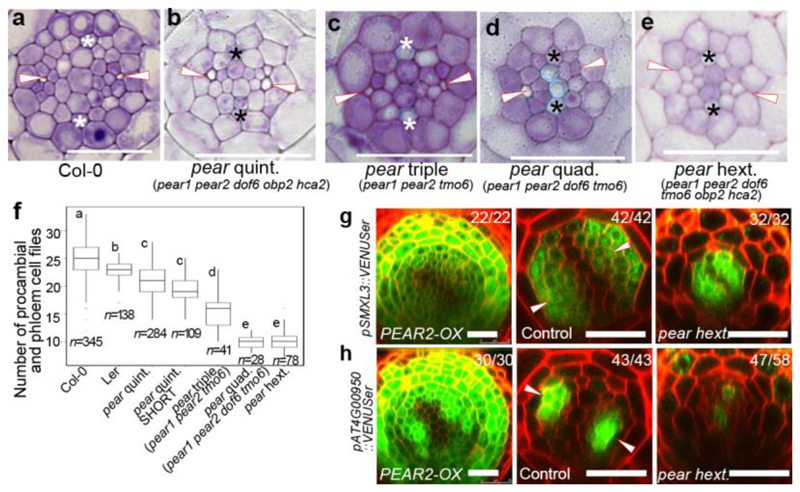
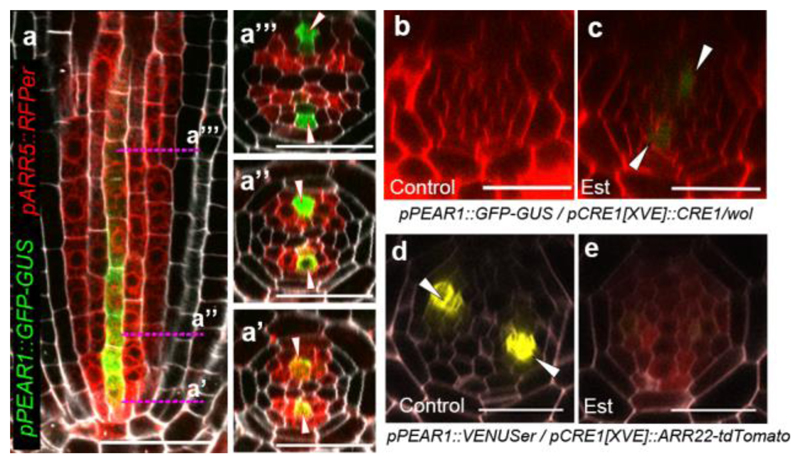
Apical growth in plants initiates upon seed germination, whereas radial growth is primed only during early ontogenesis in procambium cells and activated later by the vascular cambium. Although it is not known how radial growth is organized and regulated in plants, this system resembles the developmental competence observed in some animal systems, in which pre-existing patterns of developmental potential are established early on. Here we show that in Arabidopsis the initiation of radial growth occurs around early protophloem-sieve-element cell files of the root procambial tissue. In this domain, cytokinin signalling promotes the expression of a pair of mobile transcription factors-PHLOEM EARLY DOF 1 (PEAR1) and PHLOEM EARLY DOF 2 (PEAR2)-and their four homologues (DOF6, TMO6, OBP2 and HCA2), which we collectively name PEAR proteins. The PEAR proteins form a short-range concentration gradient that peaks at protophloem sieve elements, and activates gene expression that promotes radial growth. The expression and function of PEAR proteins are antagonized by the HD-ZIP III proteins, well-known polarity transcription factors-the expression of which is concentrated in the more-internal domain of radially non-dividing procambial cells by the function of auxin, and mobile miR165 and miR166 microRNAs. The PEAR proteins locally promote transcription of their inhibitory HD-ZIP III genes, and thereby establish a negative-feedback loop that forms a robust boundary that demarks the zone of cell division. Taken together, our data establish that during root procambial development there exists a network in which a module that links PEAR and HD-ZIP III transcription factors integrates spatial information of the hormonal domains and miRNA gradients to provide adjacent zones of dividing and more-quiescent cells, which forms a foundation for further radial growth.
植物的顶端生长始于种子萌发,而径向生长仅在原形成层细胞的早期发生,并由维管形成层 later 激活。虽然尚不清楚植物中的径向生长是如何组织和调控的,但这个系统类似于在一些动物系统中观察到的发育能力,其中预先存在的发育潜力模式很早就建立了。在这里,我们表明在拟南芥中,径向生长的起始发生在根原形成层组织的早期原生韧皮部筛分子细胞列周围。在这个区域,细胞分裂素信号促进一对移动转录因子-韧皮部早期 DOF1(PEAR1)和韧皮部早期 DOF2(PEAR2)及其四个同源物(DOF6、TMO6、OBP2 和 HCA2)的表达,我们将它们统称为 PEAR 蛋白。PEAR 蛋白形成一个短程浓度梯度,在原生韧皮部筛分子处达到峰值,并激活促进径向生长的基因表达。PEAR 蛋白的表达和功能受到 HD-ZIP III 蛋白的拮抗,HD-ZIP III 蛋白是众所周知的极性转录因子-其表达通过生长素的作用集中在径向非分裂原形成层细胞的更内部区域,并通过移动的 miR165 和 miR166 microRNAs。PEAR 蛋白局部促进其抑制性 HD-ZIP III 基因的转录,从而建立一个负反馈回路,形成一个坚固的边界,标志着细胞分裂的区域。总之,我们的数据表明,在根原形成层发育过程中,存在一个网络,该网络将连接 PEAR 和 HD-ZIP III 转录因子的模块整合到激素域和 miRNA 梯度的空间信息中,为分裂和更静止细胞的相邻区域提供基础,为进一步的径向生长奠定基础。