Institute of Human Virology, Sun Yat-sen University, Guangzhou, China.
Key Laboratory of Tropical Disease Control of Ministry of Education, Zhongshan School of Medicine, Sun Yat-sen University, Guangzhou, China.
Elife. 2019 Jan 17;8:e42426. doi: 10.7554/eLife.42426.
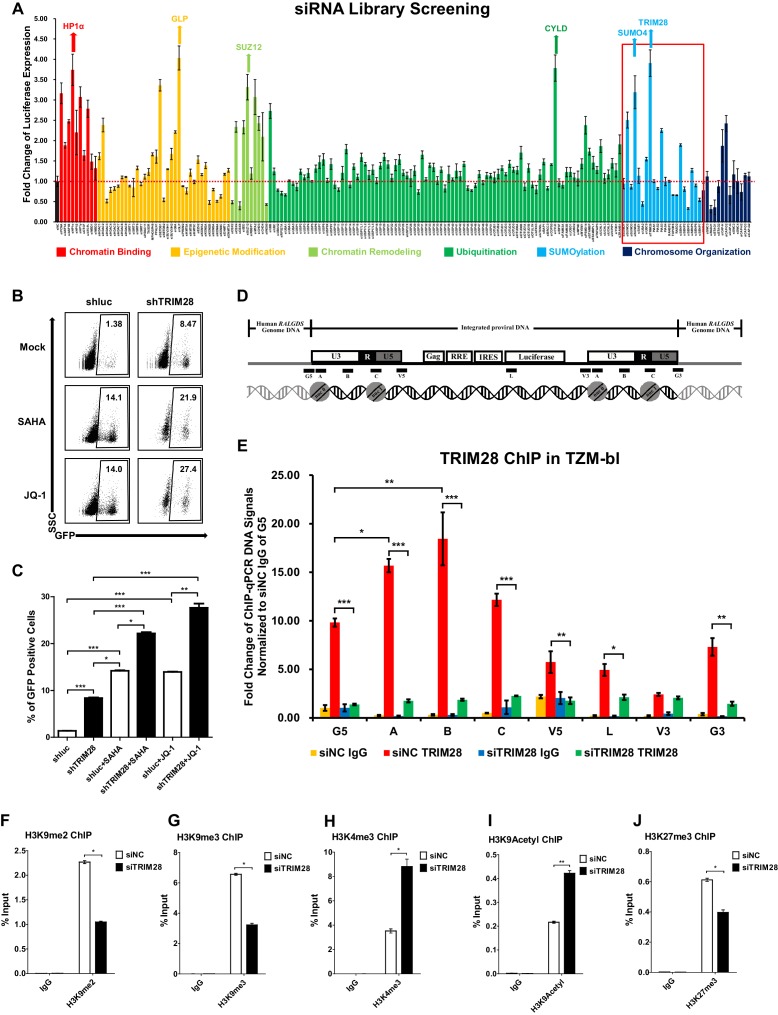
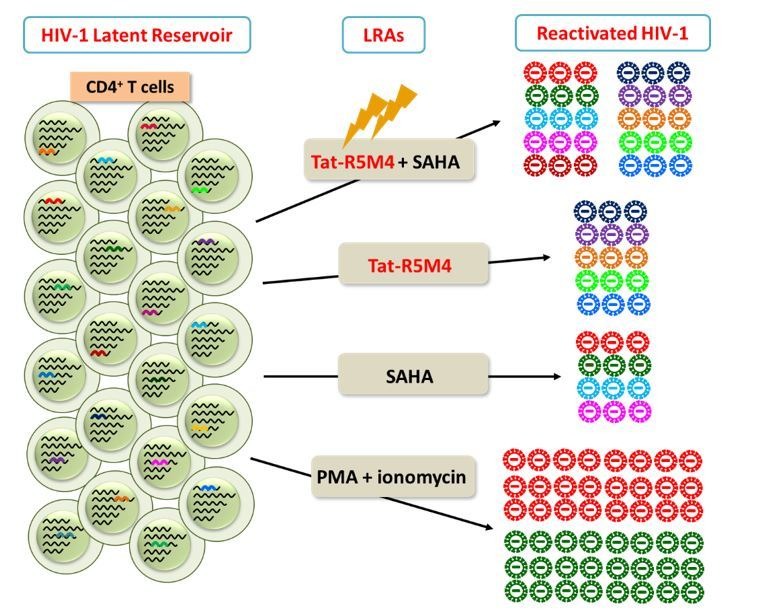
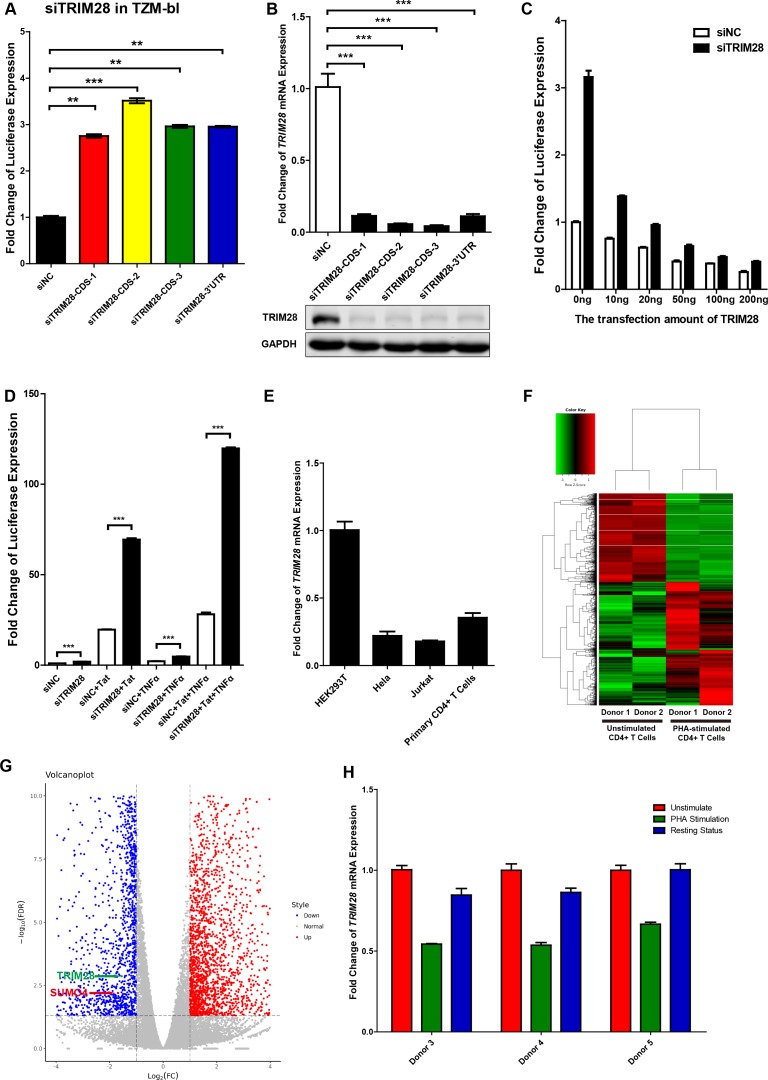
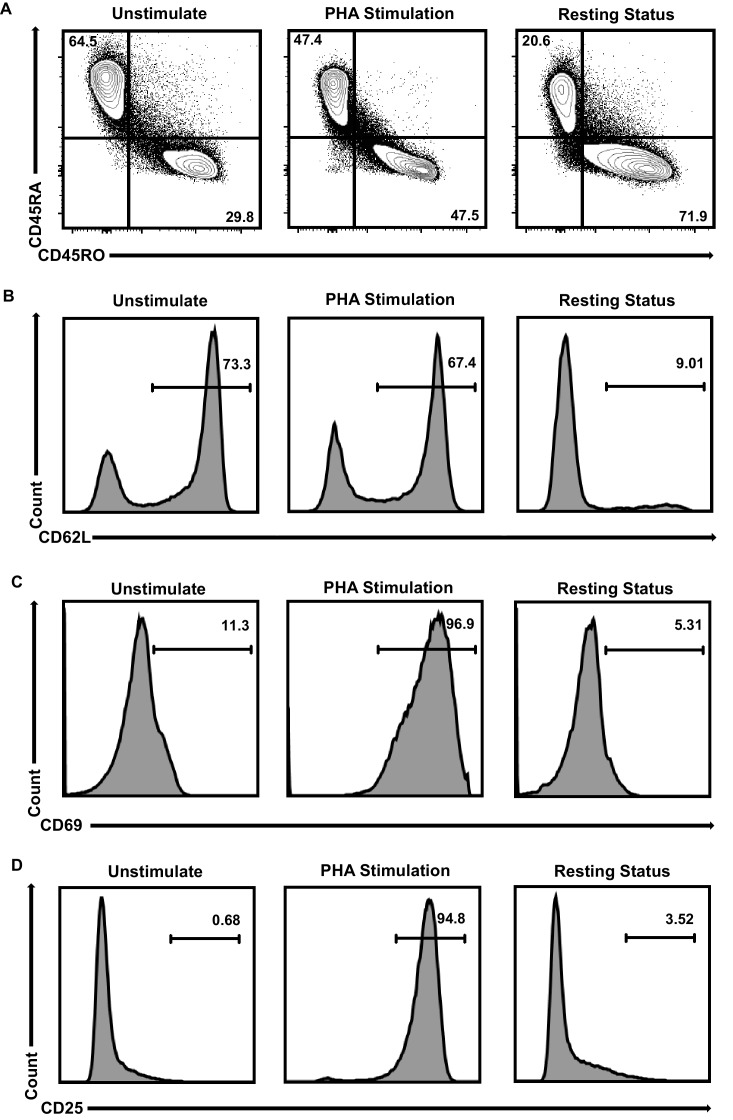
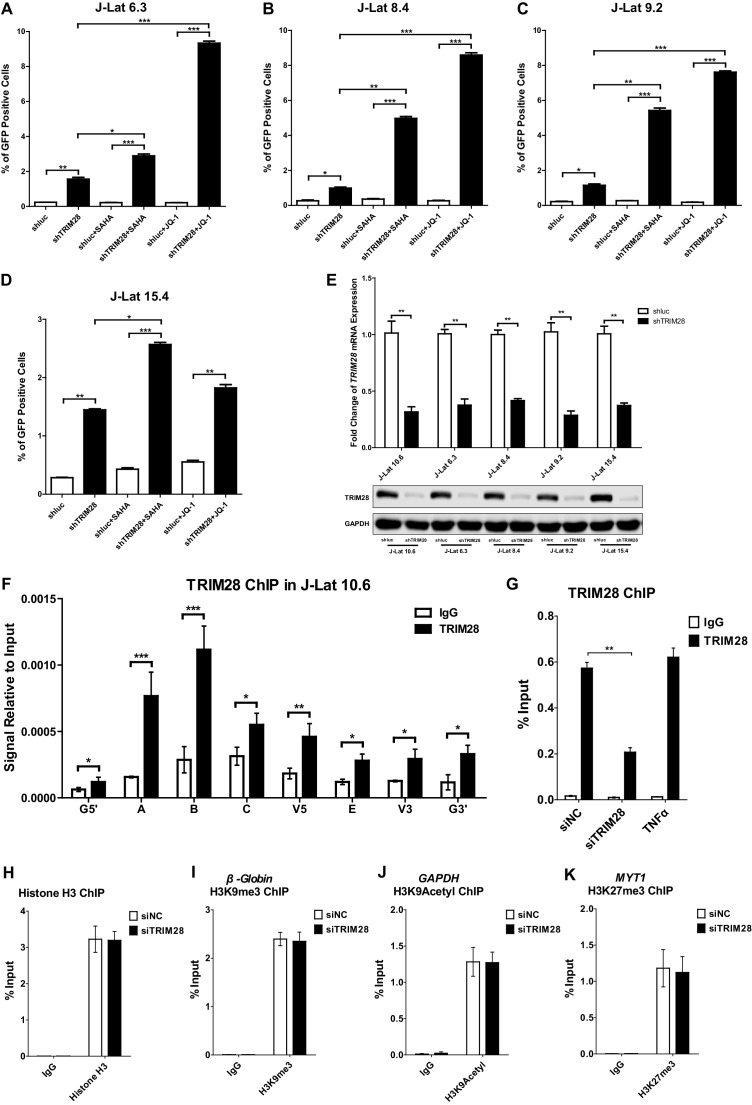
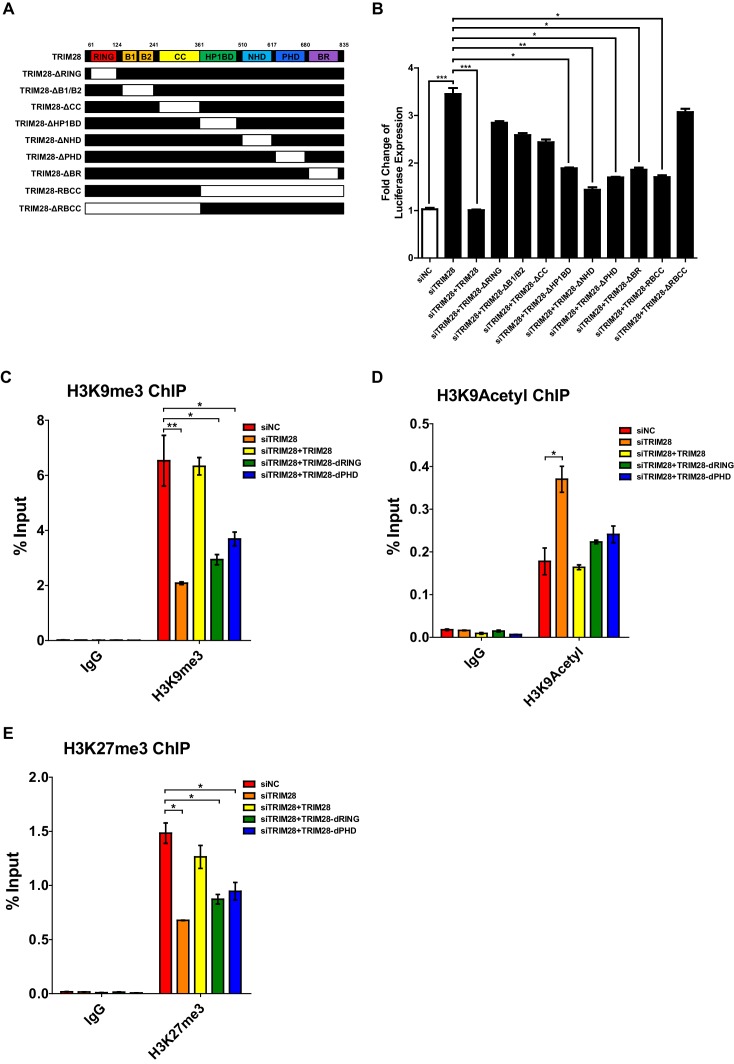
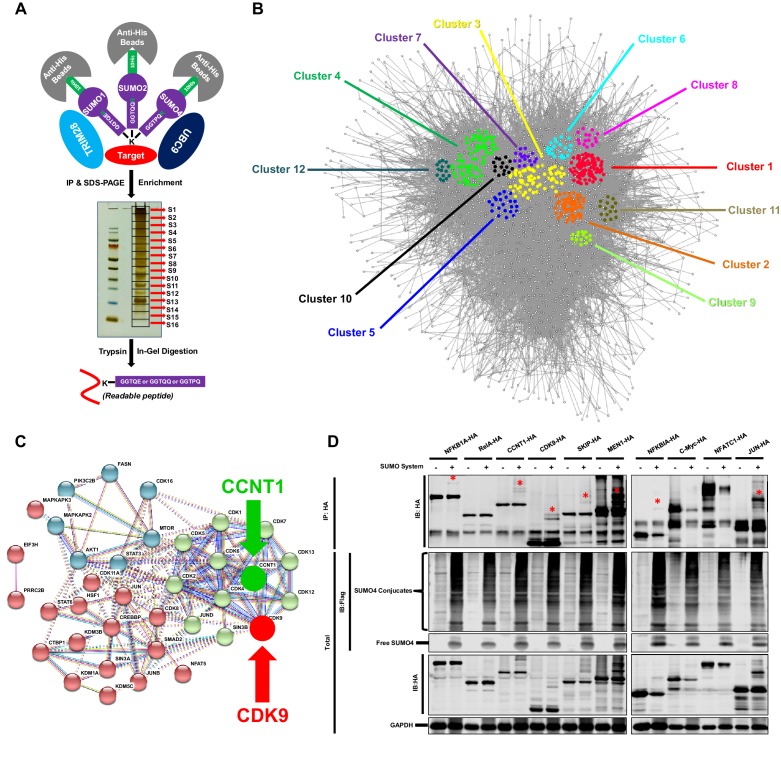
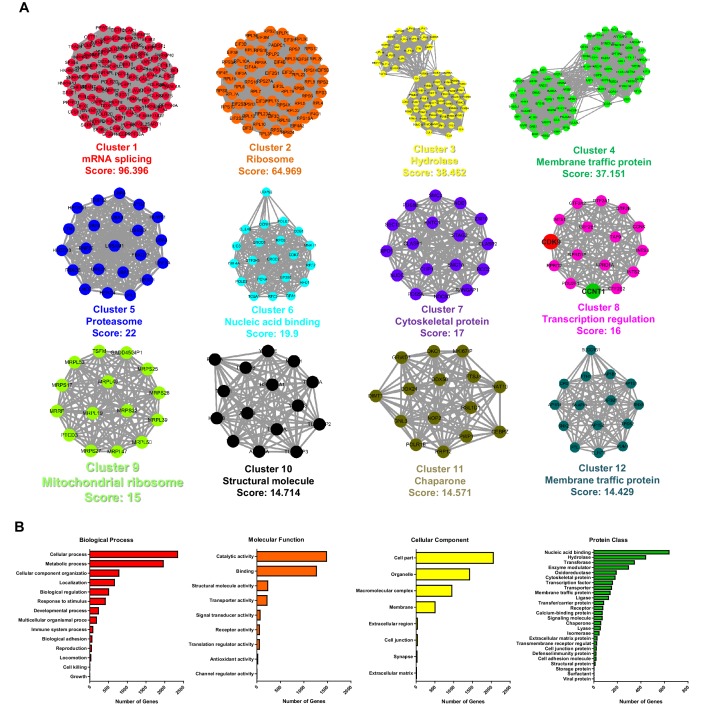
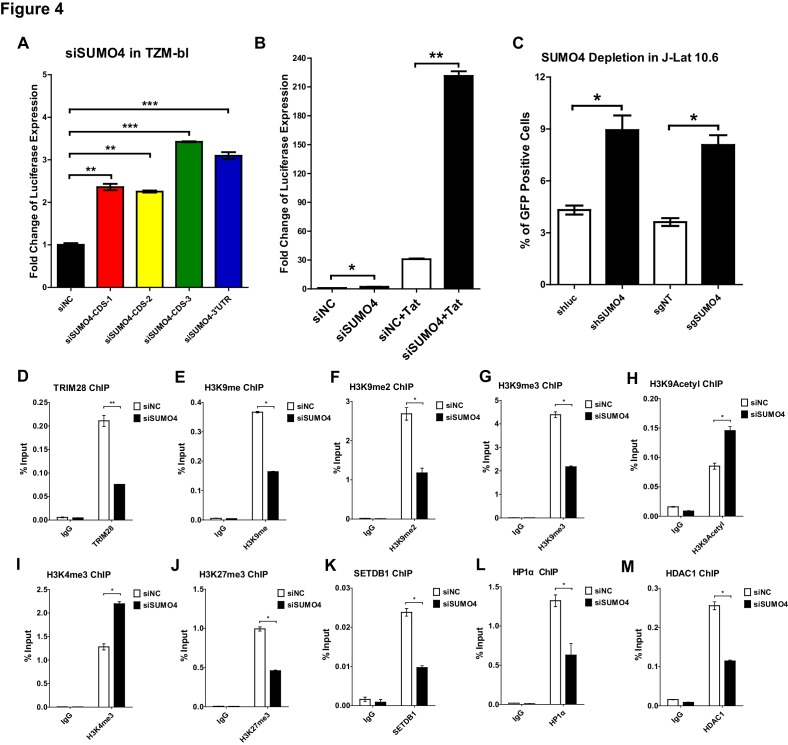
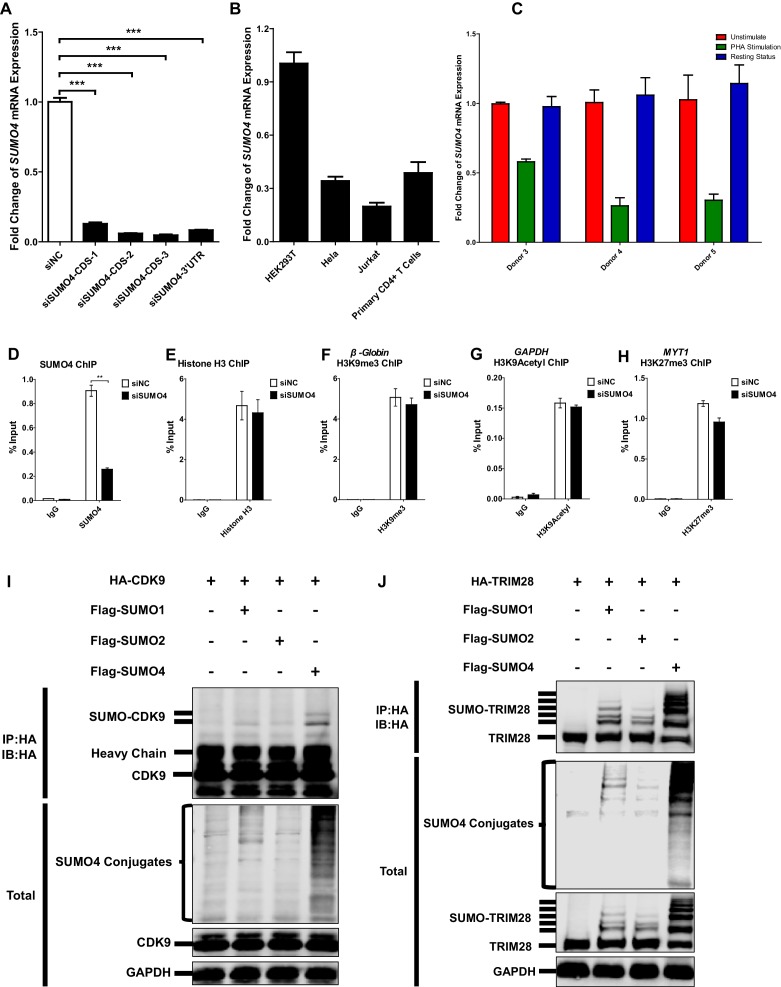
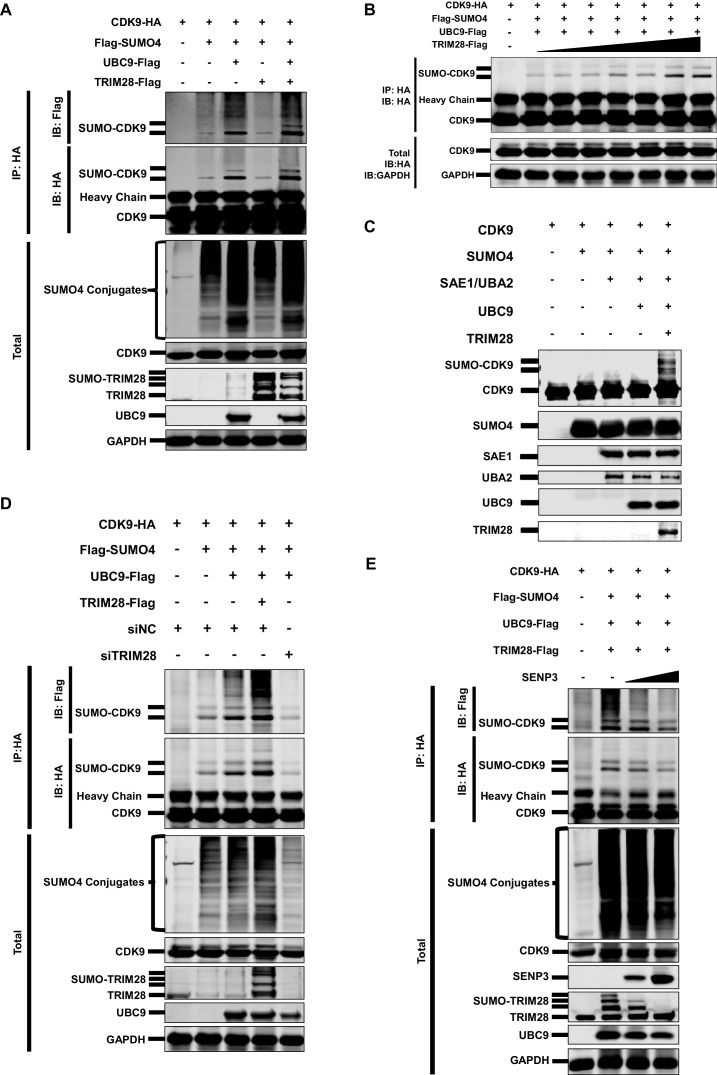
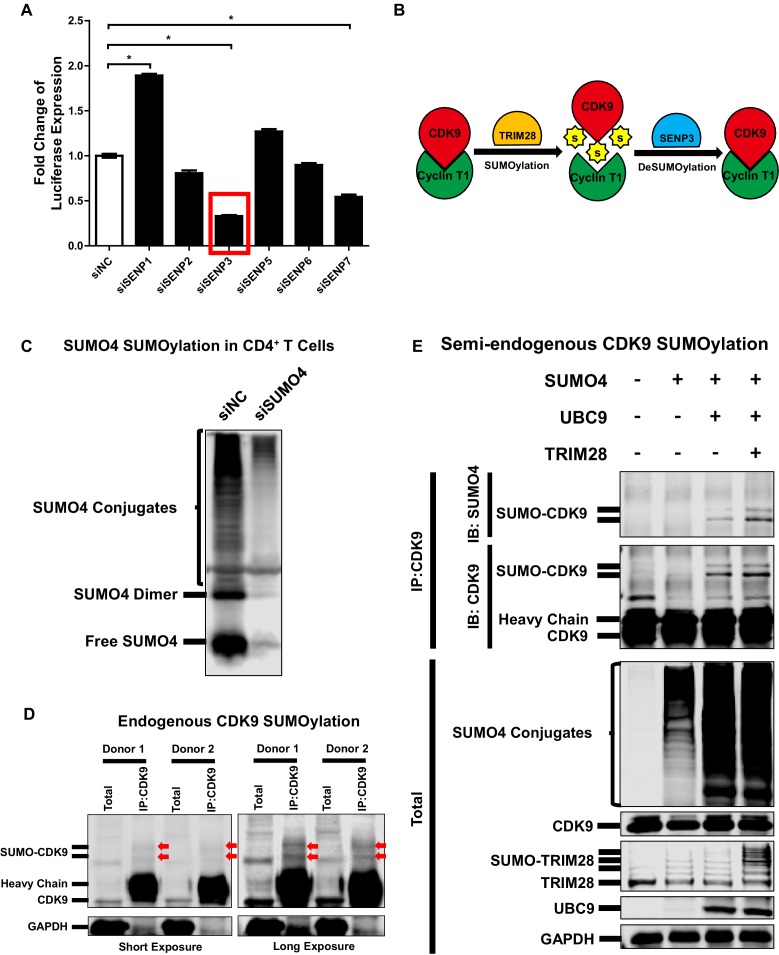
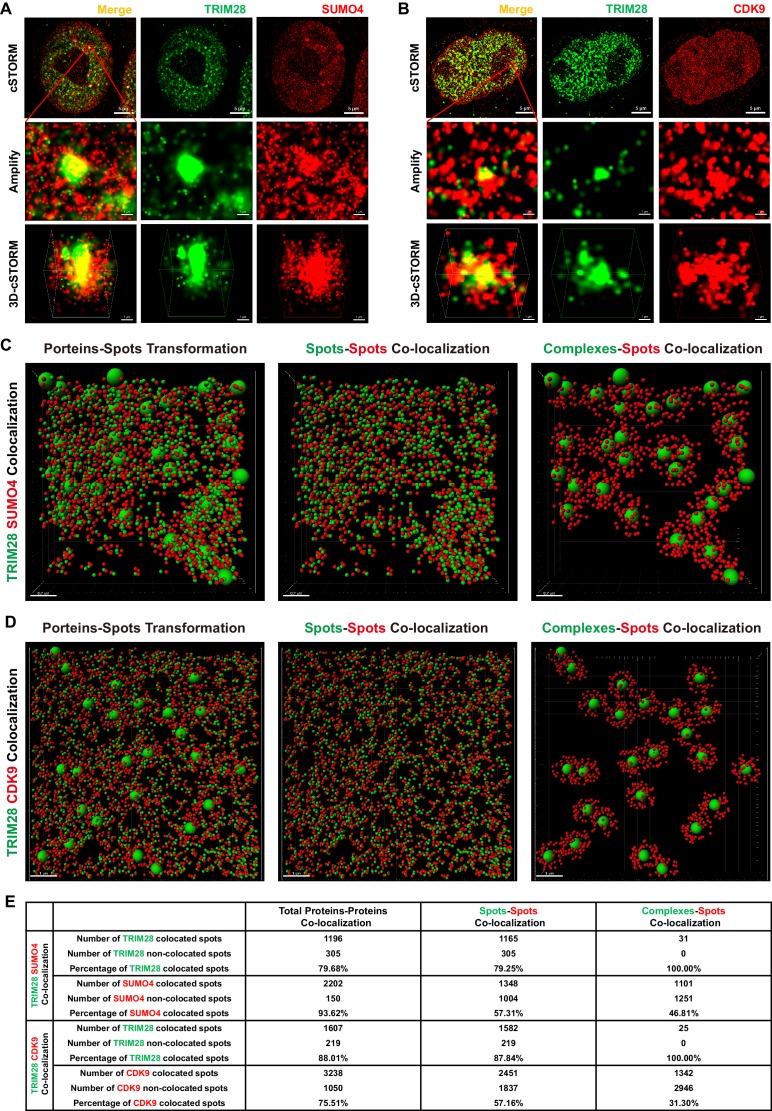
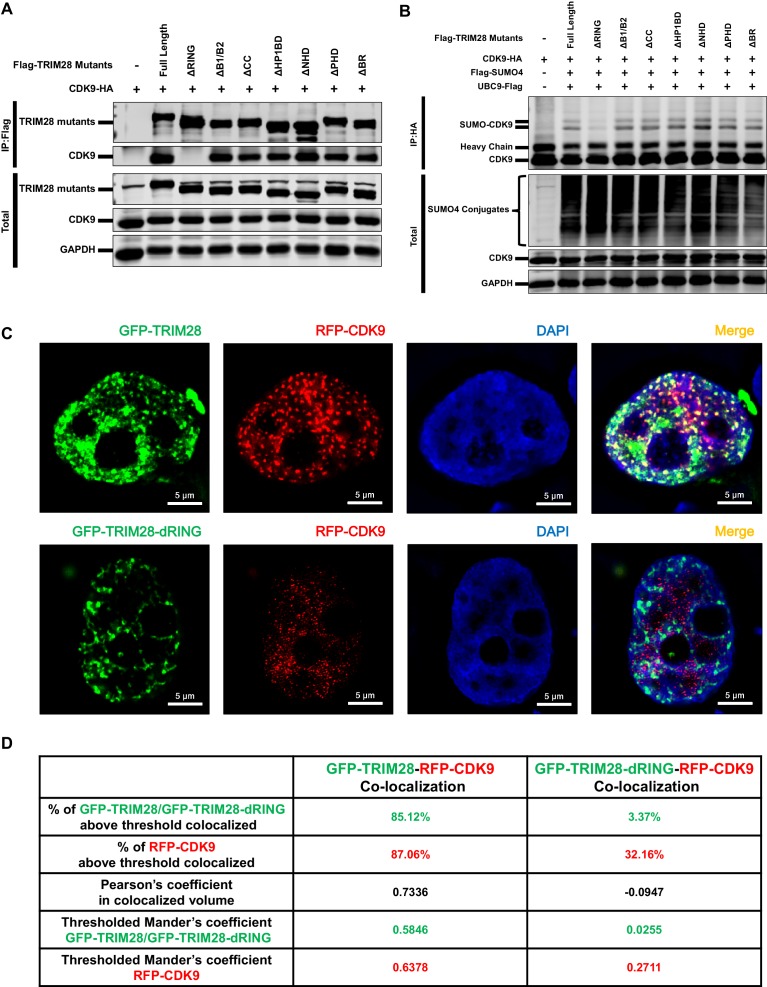
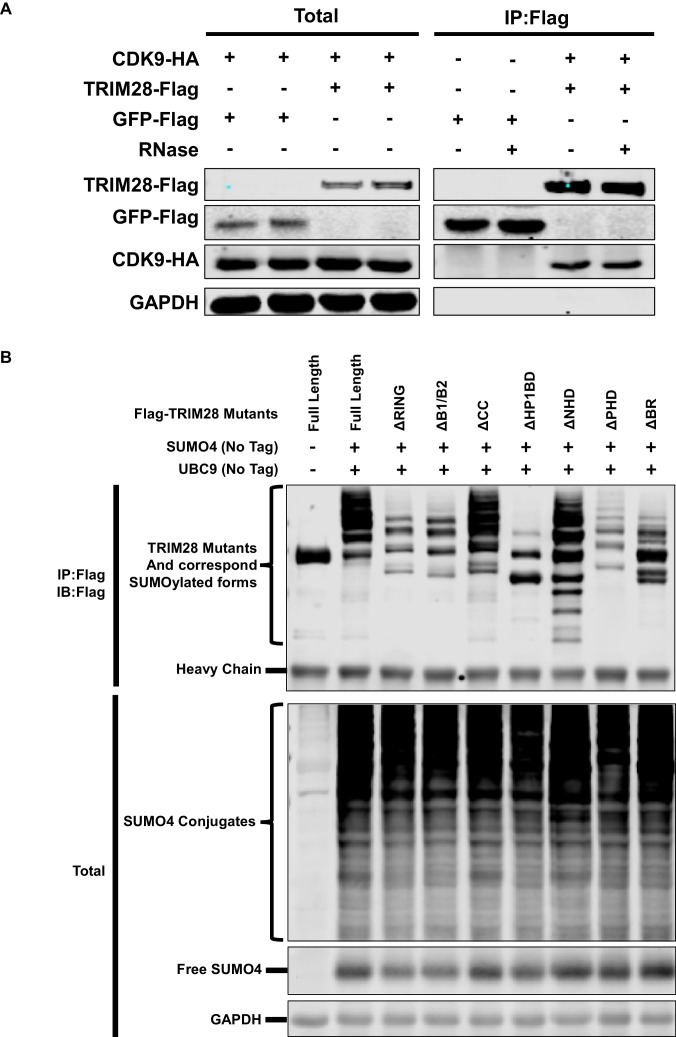
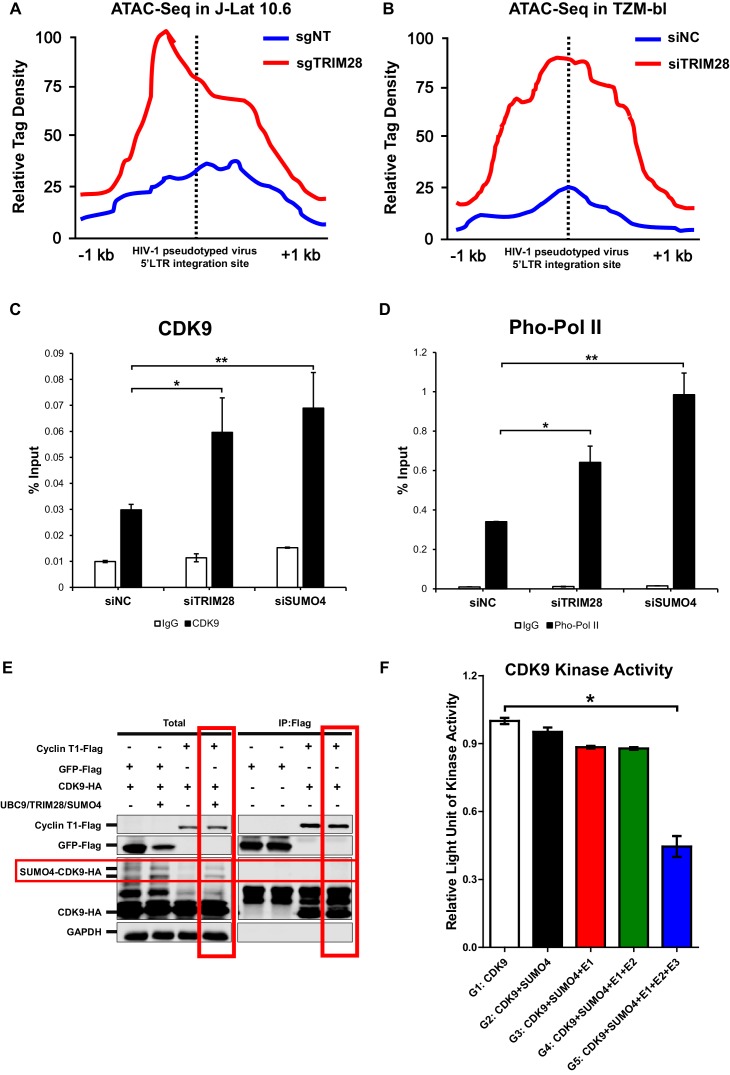
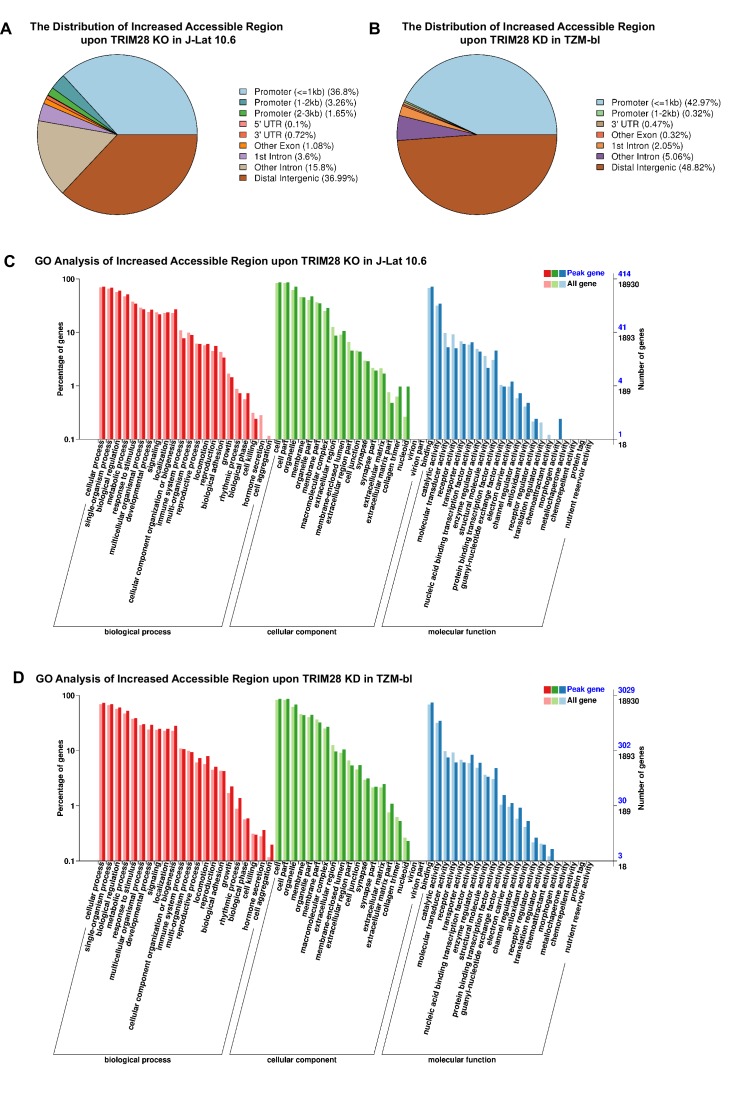
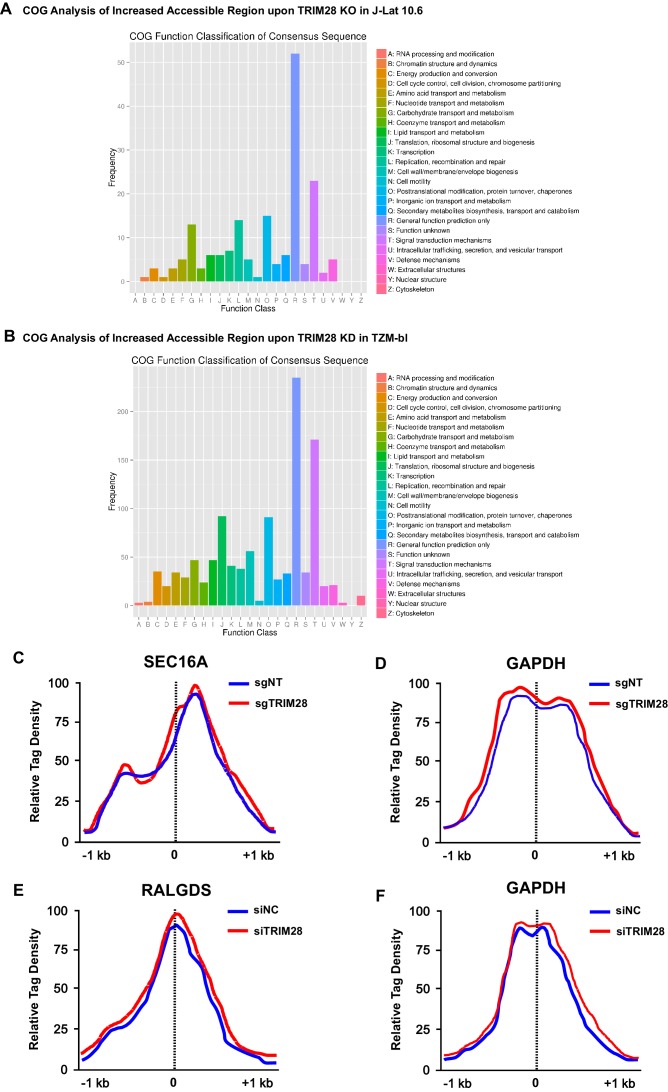
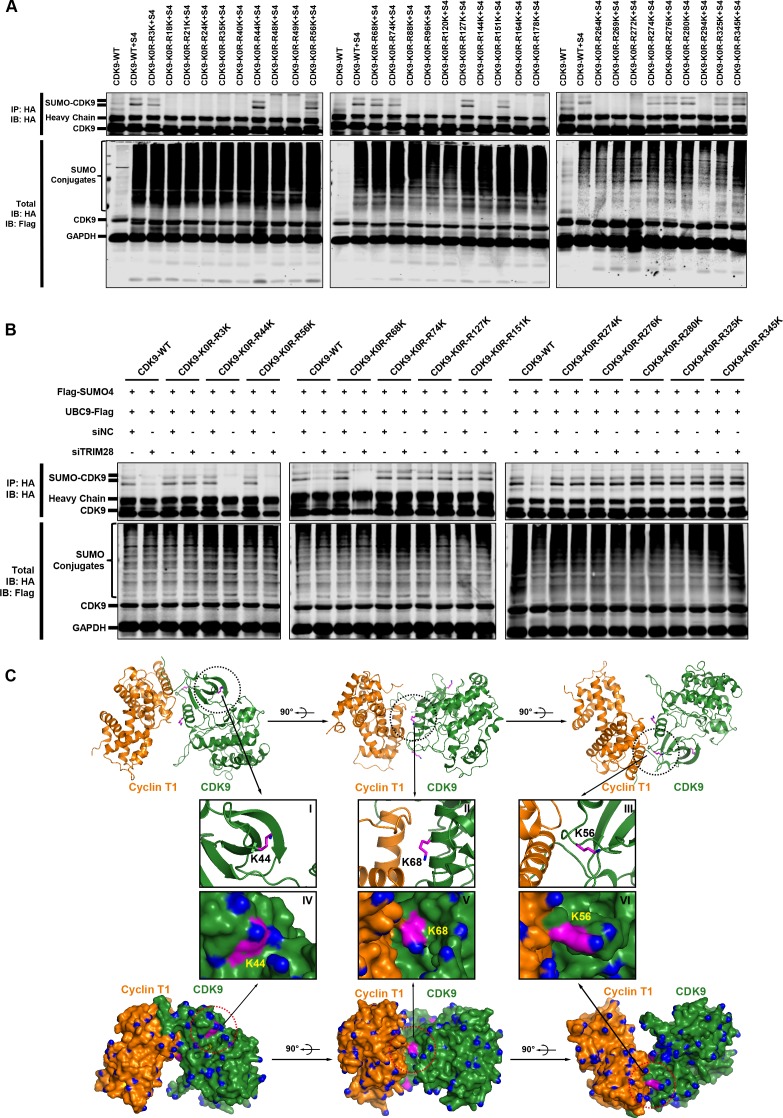
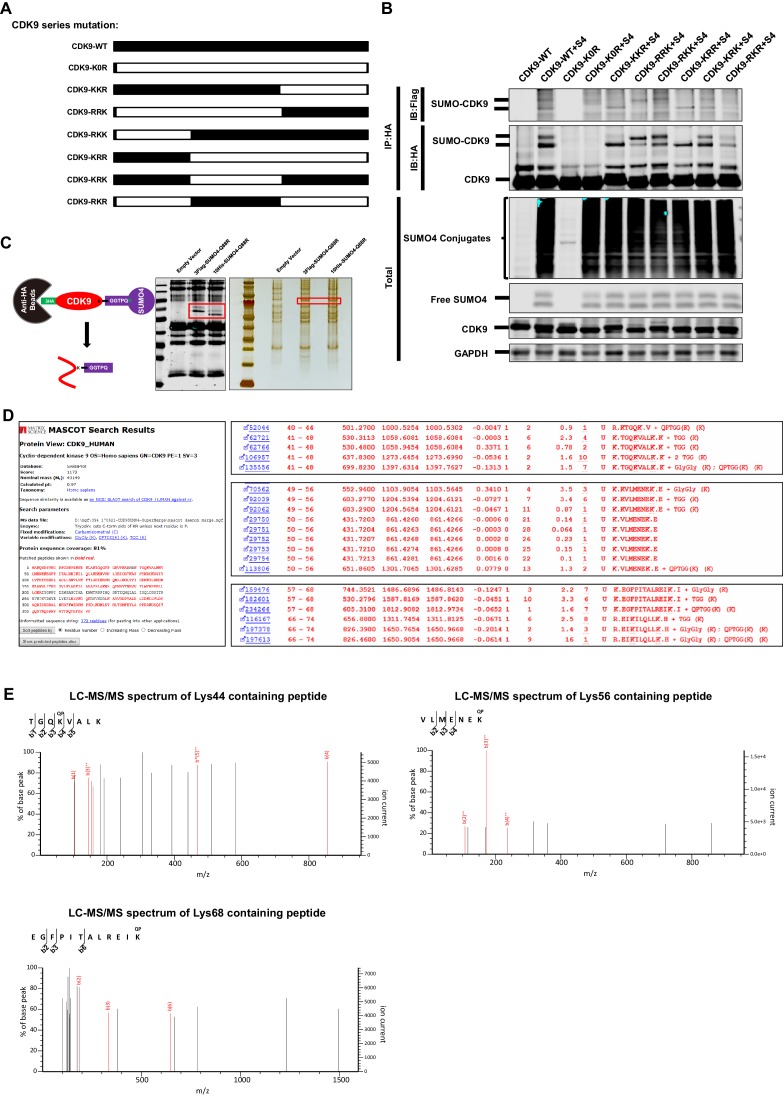
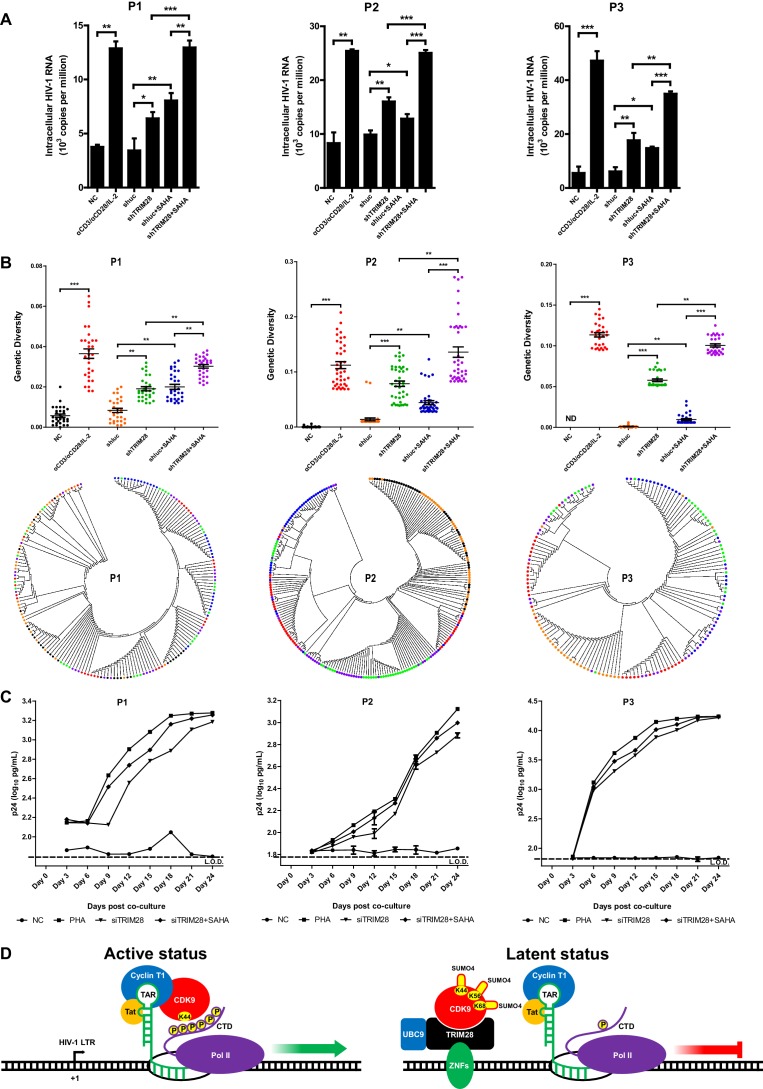
Comprehensively elucidating the molecular mechanisms of human immunodeficiency virus type 1 (HIV-1) latency is a priority to achieve a functional cure. As current 'shock' agents failed to efficiently reactivate the latent reservoir, it is important to discover new targets for developing more efficient latency-reversing agents (LRAs). Here, we found that TRIM28 potently suppresses HIV-1 expression by utilizing both SUMO E3 ligase activity and epigenetic adaptor function. Through global site-specific SUMO-MS study and serial SUMOylation assays, we identified that P-TEFb catalytic subunit CDK9 is significantly SUMOylated by TRIM28 with SUMO4. The Lys44, Lys56 and Lys68 residues on CDK9 are SUMOylated by TRIM28, which inhibits CDK9 kinase activity or prevents P-TEFb assembly by directly blocking the interaction between CDK9 and Cyclin T1, subsequently inhibits viral transcription and contributes to HIV-1 latency. The manipulation of TRIM28 and its consequent SUMOylation pathway could be the target for developing LRAs.
全面阐明人类免疫缺陷病毒 1 型(HIV-1)潜伏的分子机制是实现功能性治愈的首要任务。由于目前的“休克”剂未能有效地重新激活潜伏库,因此发现开发更有效的潜伏逆转剂(LRA)的新靶点非常重要。在这里,我们发现 TRIM28 通过利用 SUMO E3 连接酶活性和表观遗传衔接功能,有效地抑制 HIV-1 的表达。通过全局特异性 SUMO-MS 研究和连续 SUMOylation 测定,我们鉴定出 TRIM28 显著 SUMOylated P-TEFb 催化亚基 CDK9,用 SUMO4。TRIM28 对 CDK9 的 Lys44、Lys56 和 Lys68 残基进行 SUMOylation,抑制 CDK9 激酶活性或通过直接阻断 CDK9 和 Cyclin T1 之间的相互作用阻止 P-TEFb 组装,从而抑制病毒转录并有助于 HIV-1 潜伏。TRIM28 的操作及其随后的 SUMOylation 途径可能是开发 LRA 的靶点。