Nwaogu Chima J, Tieleman B Irene, Bitrus Kwanye, Cresswell Will
1Groningen Institute for Evolutionary Life Sciences, University of Groningen, P.O. Box 11103, 9700 CC Groningen, The Netherlands.
2School of Biology, University of St Andrews, Harold Mitchell Building, St Andrews, Fife KY16 9TH UK.
J Ornithol. 2018;159(4):1053-1062. doi: 10.1007/s10336-018-1574-8. Epub 2018 Jun 27.
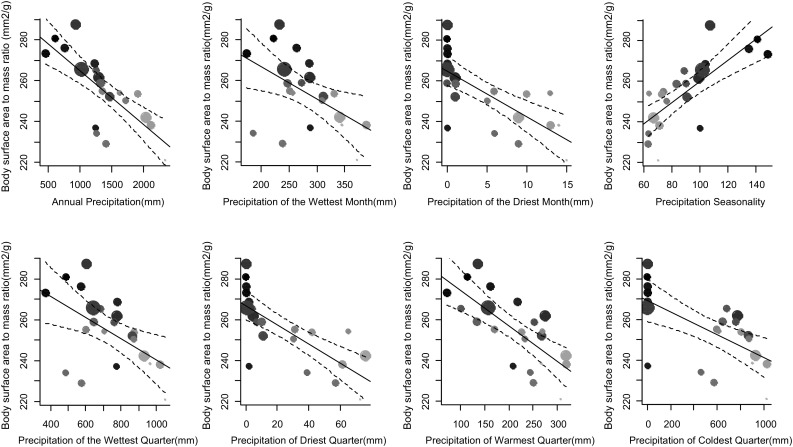
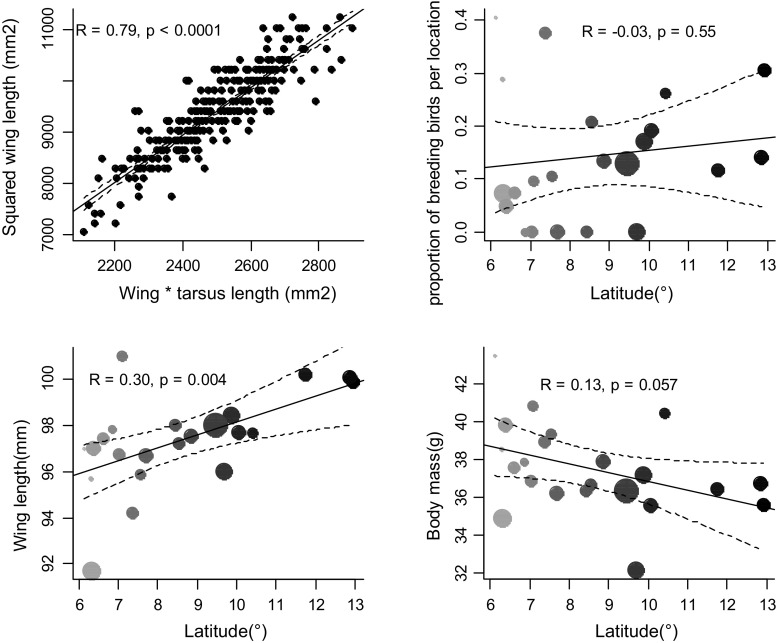
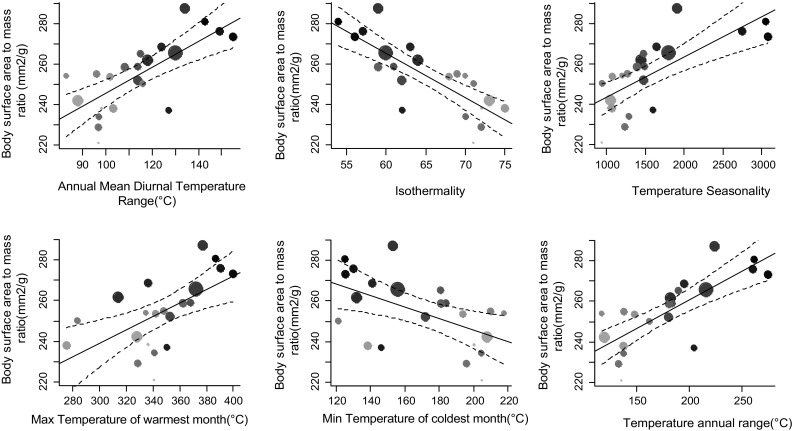
Bergmann's rule, defined as the tendency for endotherms to be larger in colder environments, is a biophysical generalization of body size variation that is frequently tested along latitudinal gradients, even though latitude is only a proxy for temperature variation. We test whether variation in temperature and aridity determine avian body size conformity to Bergmann's rule independent of latitude differences, using the ubiquitous Common Bulbul , along a West African environmental gradient. We trapped 538 birds in 22 locations between latitudes 6 and 13°N in Nigeria, and estimated average body surface area to mass ratio per location. We then modelled body surface to mass ratio using general linear models, with latitude, altitude and one of 19 bioclimatic variables extracted from http://www.worldclim.org/bioclim as predictors. We sequentially dropped latitude and altitude from each model to obtain the of the resultant models. Finally, we compared the of univariate models, where bioclimatic variables predicted body surface area to mass ratio significantly (14 out of 19), to multivariate models including latitude, altitude and a bioclimatic variable, using the Wilcoxon matched pairs test. We found that multivariate models did not perform better than univariate models with only bioclimatic variables. Six temperature and eight precipitation variables significantly predicted variation in body surface area to mass ratio between locations; in fact, 50% (seven out of 14) of these better explained variation in body surface area to mass ratio than the multivariate models. Birds showed a larger body surface area relative to body mass ratio in hotter environments independent of latitude or altitude, which conforms to Bergmann's rule. Yet, a combination of morphometric analyses and controlled temperature-exposure experiments is required to prove the proposed relationship between relative body surface area and thermoregulation in endotherms.
伯格曼法则定义为恒温动物在寒冷环境中体型趋于更大的趋势,它是对体型变化的一种生物物理学概括,尽管纬度只是温度变化的一个替代指标,但该法则经常沿着纬度梯度进行检验。我们利用广泛分布的家八哥,沿着西非环境梯度,测试温度和干旱度的变化是否独立于纬度差异,决定鸟类体型是否符合伯格曼法则。我们在尼日利亚北纬6度至13度之间的22个地点捕获了538只鸟,并估算每个地点的平均体表面积与质量比。然后,我们使用一般线性模型,以纬度、海拔以及从http://www.worldclim.org/bioclim提取的19个生物气候变量之一作为预测因子,对体表面积与质量比进行建模。我们从每个模型中依次剔除纬度和海拔,以获得所得模型的结果。最后,我们使用威尔科克森配对检验,比较单变量模型(其中19个生物气候变量中有14个显著预测了体表面积与质量比)与包含纬度、海拔和一个生物气候变量的多变量模型的结果。我们发现,多变量模型的表现并不优于仅包含生物气候变量的单变量模型。六个温度变量和八个降水变量显著预测了不同地点之间体表面积与质量比的变化;事实上,其中50%(14个中的7个)比多变量模型能更好地解释体表面积与质量比的变化。无论纬度或海拔如何,鸟类在较热环境中的体表面积与体重比相对更大,这符合伯格曼法则。然而,需要结合形态计量分析和控制温度暴露实验,来证明恒温动物相对体表面积与体温调节之间的拟议关系。