Centre for Gene Regulation and Expression, School of Life Sciences, University of Dundee, Dow Street, Dundee DD1 5EH, UK.
Centre for Gene Regulation and Expression, School of Life Sciences, University of Dundee, Dow Street, Dundee DD1 5EH, UK.
Curr Biol. 2019 May 6;29(9):1536-1544.e4. doi: 10.1016/j.cub.2019.03.051. Epub 2019 Apr 18.
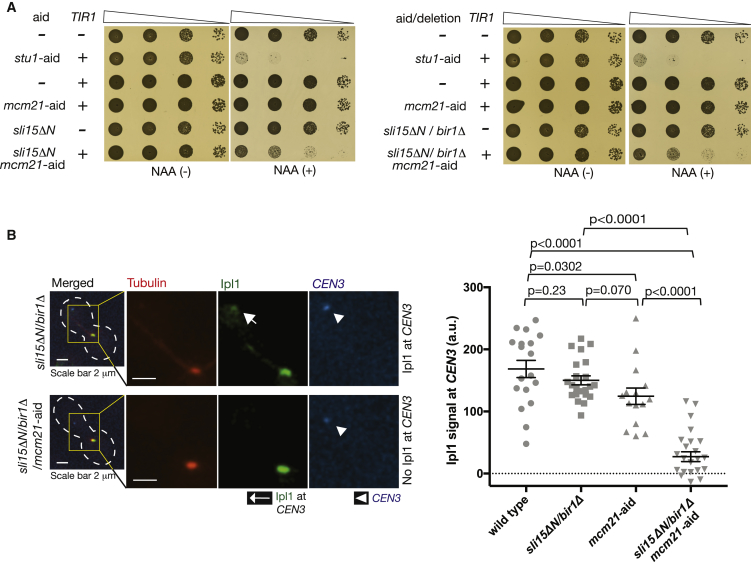
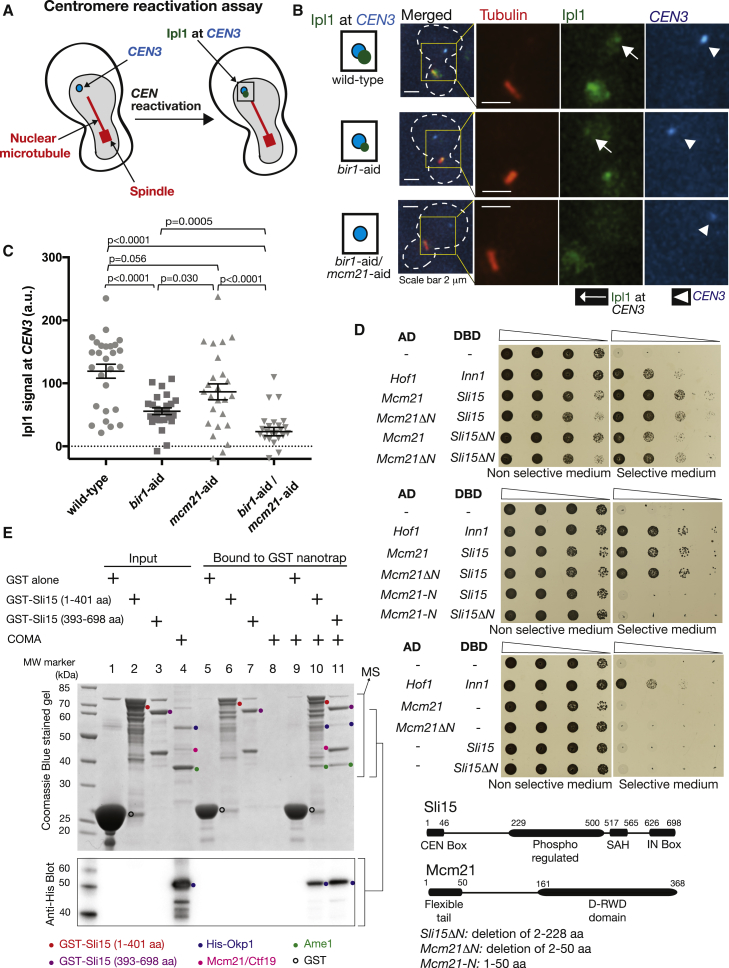
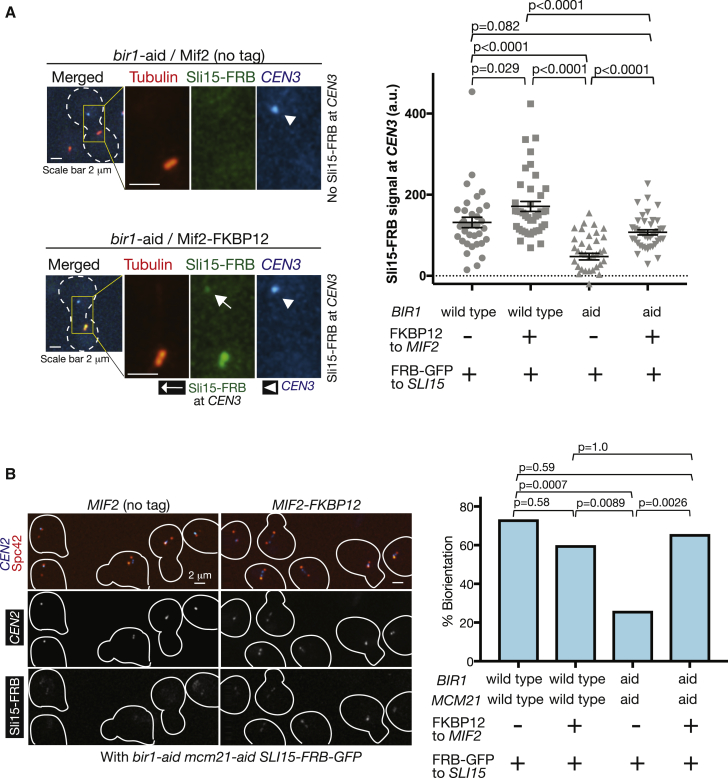
For proper chromosome segregation in mitosis, sister kinetochores must interact with microtubules from opposite spindle poles (chromosome bi-orientation) [1, 2]. To promote bi-orientation, Aurora B kinase disrupts aberrant kinetochore-microtubule interactions [3-6]. It has long been debated how Aurora B halts this action when bi-orientation is established and tension is applied across sister kinetochores. A popular explanation for it is that, upon bi-orientation, sister kinetochores are pulled in opposite directions, stretching the outer kinetochores [7, 8] and moving Aurora B substrates away from Aurora-B-localizing sites at centromeres (spatial separation model) [3, 5, 9]. This model predicts that Aurora B localization at centromeres is required for bi-orientation. However, this notion was challenged by the observation that Bir1 (yeast survivin), which recruits Ipl1-Sli15 (yeast Aurora B-INCENP) to centromeres, can become dispensable for bi-orientation [10]. This raised the possibility that Aurora B localization at centromeres is dispensable for bi-orientation. Alternatively, there might be a Bir1-independent mechanism for recruiting Ipl1-Sli15 to centromeres or inner kinetochores [5, 9]. Here, we show that the COMA inner kinetochore sub-complex physically interacts with Sli15, recruits Ipl1-Sli15 to the inner kinetochore, and promotes chromosome bi-orientation, independently of Bir1, in budding yeast. Moreover, using an engineered recruitment of Ipl1-Sli15 to the inner kinetochore when both Bir1 and COMA are defective, we show that localization of Ipl1-Sli15 at centromeres or inner kinetochores is required for bi-orientation. Our results give important insight into how Aurora B disrupts kinetochore-microtubule interaction in a tension-dependent manner to promote chromosome bi-orientation.
为了实现有丝分裂过程中染色体的正确分离,姐妹动粒必须与来自纺锤体相反两极的微管相互作用(染色体双定向)[1,2]。为了促进双定向,Aurora B 激酶破坏异常的动粒-微管相互作用[3-6]。长期以来,人们一直在争论 Aurora B 在双定向建立和姐妹动粒之间施加张力时如何停止这种作用。一个流行的解释是,双定向后,姐妹动粒被向相反方向拉动,拉伸外动粒[7,8]并将 Aurora B 底物从着丝粒处的 Aurora-B 定位位点移开(空间分离模型)[3,5,9]。该模型预测,着丝粒处的 Aurora B 定位对于双定向是必需的。然而,Bir1(酵母 survivin)的观察结果挑战了这一观点,Bir1 将 Ipl1-Sli15(酵母 Aurora B-INCENP)招募到着丝粒,在双定向中变得可有可无[10]。这就提出了 Aurora B 在着丝粒处的定位对于双定向可有可无的可能性。或者,可能存在一种独立于 Bir1 的机制将 Ipl1-Sli15 招募到着丝粒或内动粒[5,9]。在这里,我们证明了 COMA 内动粒亚复合物与 Sli15 物理相互作用,将 Ipl1-Sli15 招募到内动粒,并促进染色体双定向,独立于 Bir1,在芽殖酵母中。此外,使用工程化方法将 Ipl1-Sli15 招募到内动粒,同时使 Bir1 和 COMA 均失活,我们表明 Ipl1-Sli15 在内动粒或着丝粒处的定位对于双定向是必需的。我们的结果为 Aurora B 如何以张力依赖的方式破坏动粒-微管相互作用以促进染色体双定向提供了重要的见解。