Department of Microbiology, Biochemistry and Molecular Genetics, International Center for Public Health, New Jersey Medical School, Rutgers, The State University of New Jersey, Newark, New Jersey, United States of America.
Department of Molecular Biophysics and Biochemistry, Yale University School of Medicine, New Haven, Connecticut, United States of America.
PLoS Genet. 2019 Aug 1;15(8):e1008294. doi: 10.1371/journal.pgen.1008294. eCollection 2019 Aug.
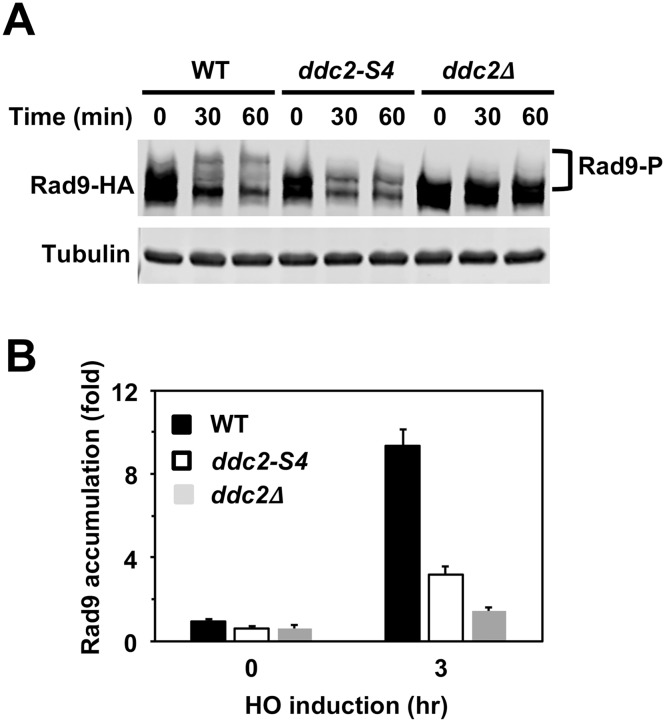
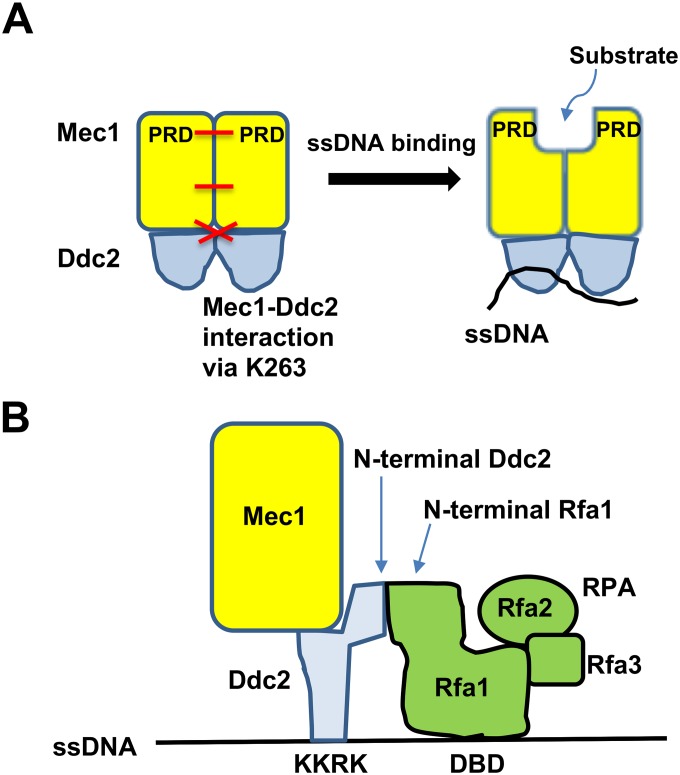
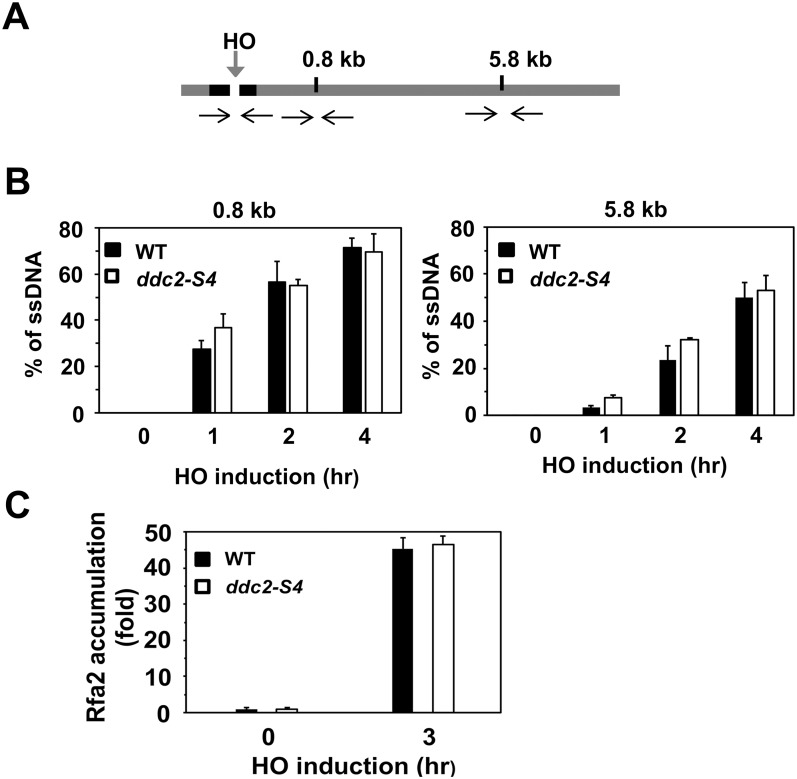
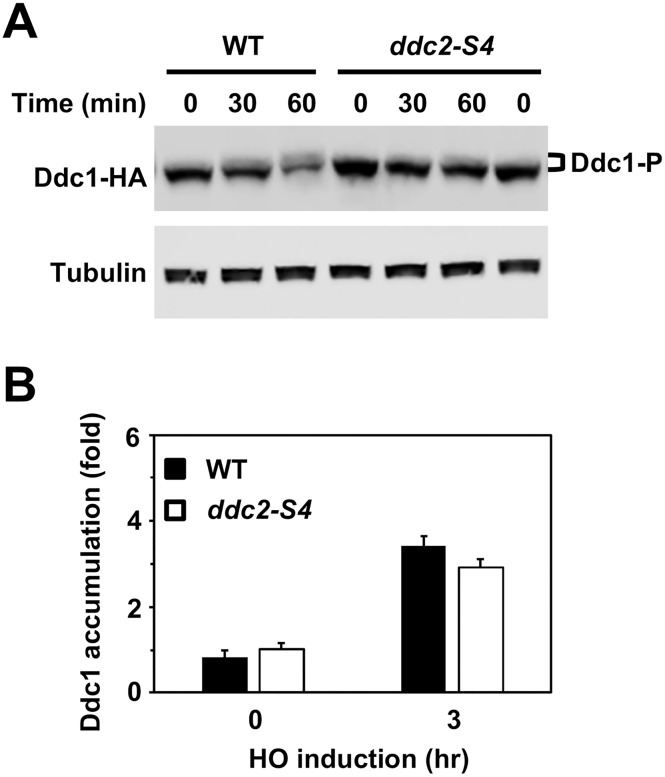
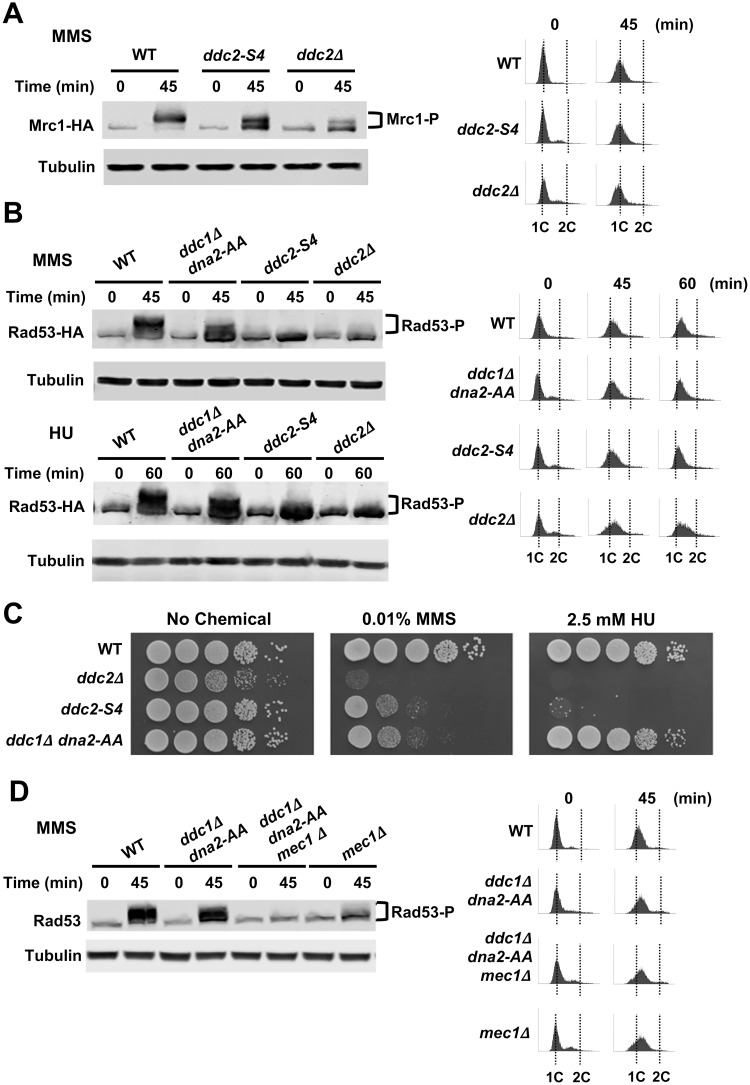
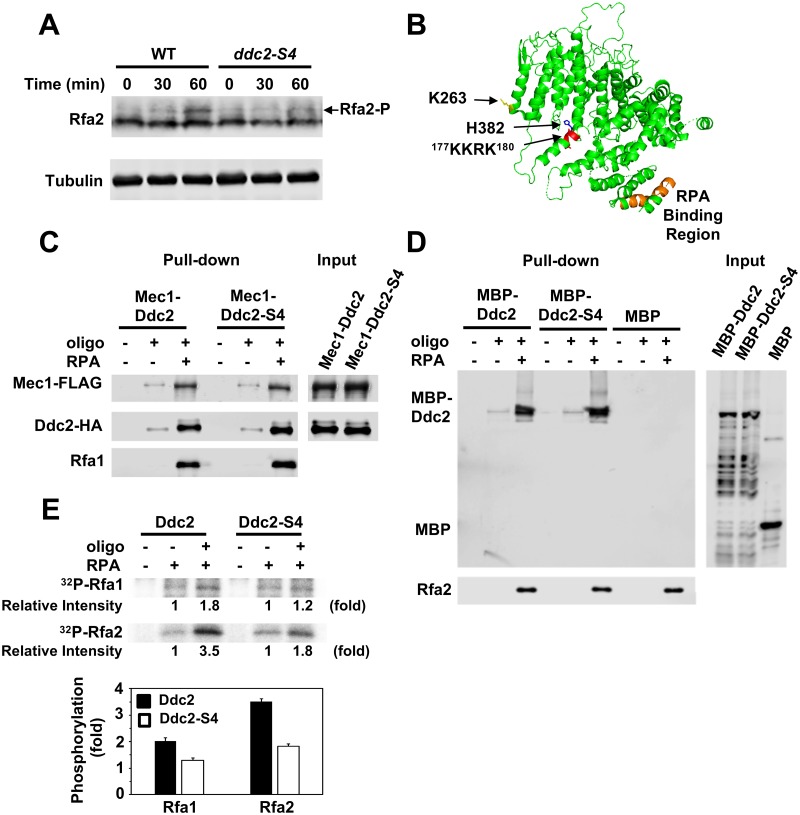
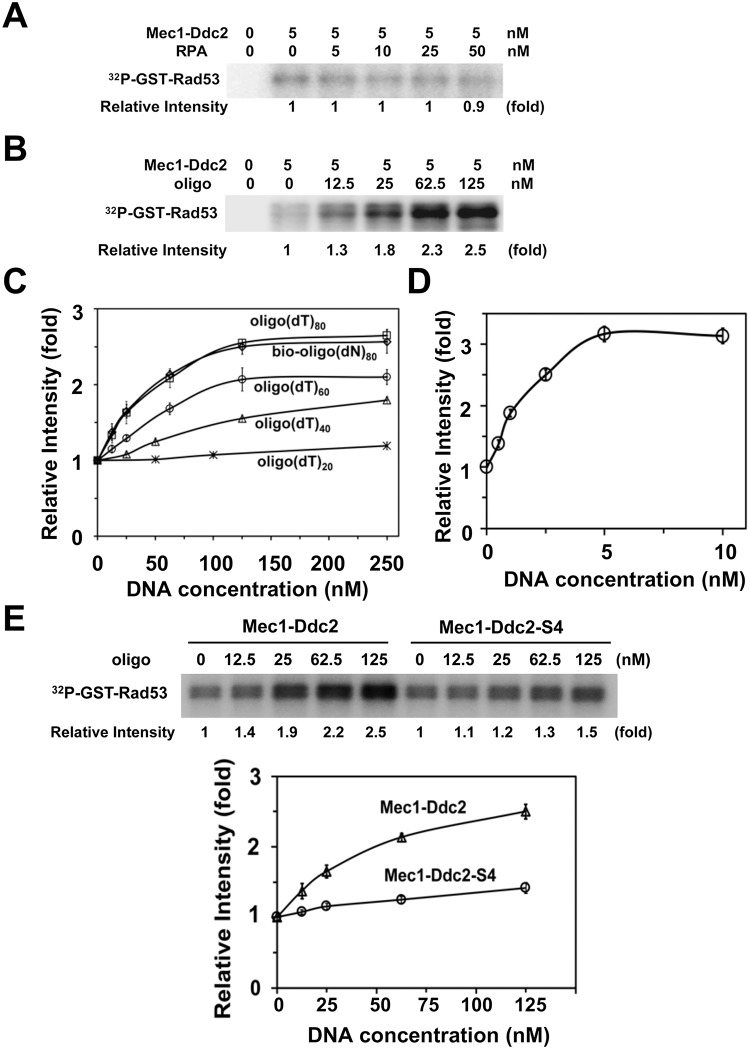
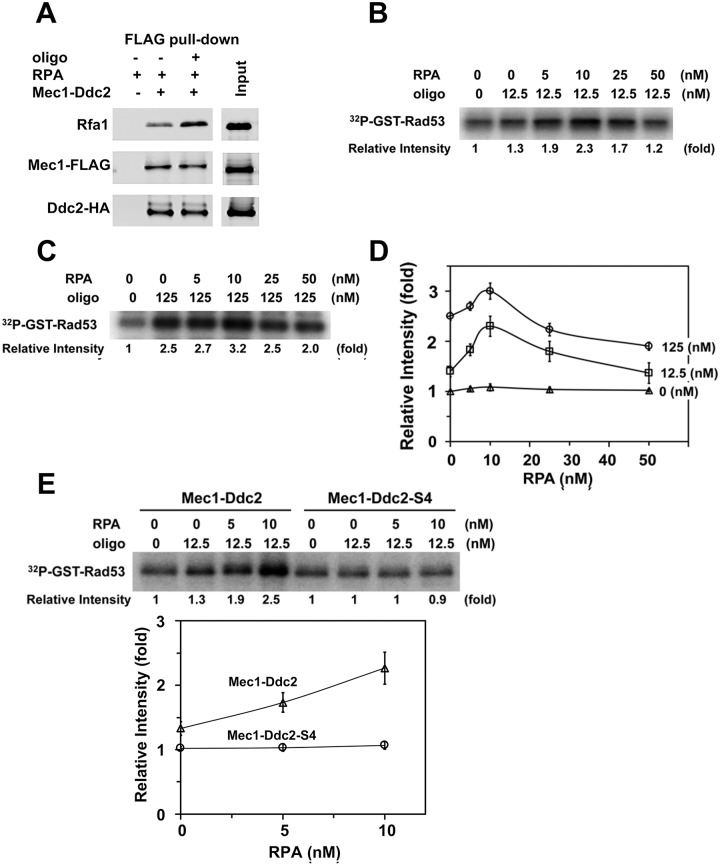
The DNA damage checkpoint response is controlled by the phosphatidylinositol 3-kinase-related kinases (PIKK), including ataxia telangiectasia-mutated (ATM) and ATM and Rad3-related (ATR). ATR forms a complex with its partner ATRIP. In budding yeast, ATR and ATRIP correspond to Mec1 and Ddc2, respectively. ATRIP/Ddc2 interacts with replication protein A-bound single-stranded DNA (RPA-ssDNA) and recruits ATR/Mec1 to sites of DNA damage. Mec1 is stimulated by the canonical activators including Ddc1, Dpb11 and Dna2. We have characterized the ddc2-S4 mutation and shown that Ddc2 not only recruits Mec1 to sites of DNA damage but also stimulates Mec1 kinase activity. However, the underlying mechanism of Ddc2-dependent Mec1 activation remains to be elucidated. Here we show that Ddc2 promotes Mec1 activation independently of Ddc1/Dpb11/Dna2 function in vivo and through ssDNA recognition in vitro. The ddc2-S4 mutation diminishes damage-induced phosphorylation of the checkpoint mediators, Rad9 and Mrc1. Rad9 controls checkpoint throughout the cell-cycle whereas Mrc1 is specifically required for the S-phase checkpoint. Notably, S-phase checkpoint signaling is more defective in ddc2-S4 mutants than in cells where the Mec1 activators (Ddc1/Dpb11 and Dna2) are dysfunctional. To understand a role of Ddc2 in Mec1 activation, we reconstituted an in vitro assay using purified Mec1-Ddc2 complex, RPA and ssDNA. Whereas ssDNA stimulates kinase activity of the Mec1-Ddc2 complex, RPA does not. However, RPA can promote ssDNA-dependent Mec1 activation. Neither ssDNA nor RPA-ssDNA efficiently stimulates the Mec1-Ddc2 complex containing Ddc2-S4 mutant. Together, our data support a model in which Ddc2 promotes Mec1 activation at RPA-ssDNA tracts.
DNA 损伤检查点反应由磷脂酰肌醇 3-激酶相关激酶(PIKK)控制,包括共济失调毛细血管扩张症突变(ATM)和 ATM 和 Rad3 相关(ATR)。ATR 与它的伴侣 ATRIP 形成复合物。在芽殖酵母中,ATR 和 ATRIP 分别对应于 Mec1 和 Ddc2。ATRIP/Ddc2 与复制蛋白 A 结合的单链 DNA(RPA-ssDNA)相互作用,并将 ATR/Mec1 招募到 DNA 损伤部位。Mec1 被包括 Ddc1、Dpb11 和 Dna2 在内的经典激活剂刺激。我们已经对 ddc2-S4 突变进行了特征描述,并表明 Ddc2 不仅将 Mec1 招募到 DNA 损伤部位,而且还刺激 Mec1 激酶活性。然而,Ddc2 依赖性 Mec1 激活的潜在机制仍有待阐明。在这里,我们表明 Ddc2 促进 Mec1 激活,而不依赖于体内的 Ddc1/Dpb11/Dna2 功能,并且通过体外的 ssDNA 识别。ddc2-S4 突变减少了损伤诱导的检查点介质 Rad9 和 Mrc1 的磷酸化。Rad9 在整个细胞周期中控制检查点,而 Mrc1 专门用于 S 期检查点。值得注意的是,ddc2-S4 突变体中的 S 期检查点信号比 Mec1 激活剂(Ddc1/Dpb11 和 Dna2)功能失调的细胞中的更有缺陷。为了了解 Ddc2 在 Mec1 激活中的作用,我们使用纯化的 Mec1-Ddc2 复合物、RPA 和 ssDNA 重新构建了体外测定。虽然 ssDNA 刺激 Mec1-Ddc2 复合物的激酶活性,但 RPA 没有。然而,RPA 可以促进 ssDNA 依赖性 Mec1 激活。ssDNA 或 RPA-ssDNA 都不能有效地刺激包含 ddc2-S4 突变体的 Mec1-Ddc2 复合物。总之,我们的数据支持一种模型,即 Ddc2 在 RPA-ssDNA 片段上促进 Mec1 激活。