Harriton High School, Rosemont, Pennsylvania, United States of America.
Department of Chemical and Biological Engineering, University of Pennsylvania, Philadelphia, Pennsylvania, United States of America.
PLoS Comput Biol. 2019 Oct 7;15(10):e1007295. doi: 10.1371/journal.pcbi.1007295. eCollection 2019 Oct.
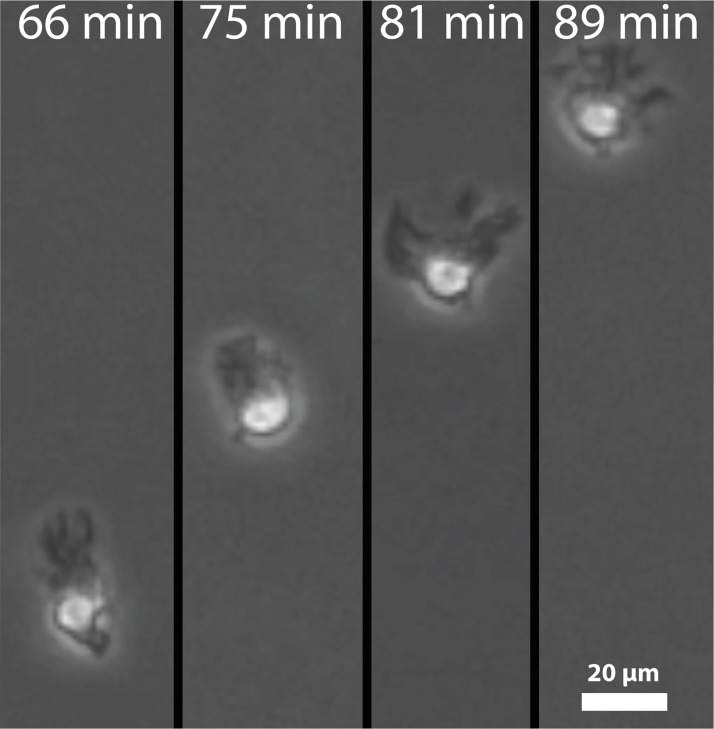
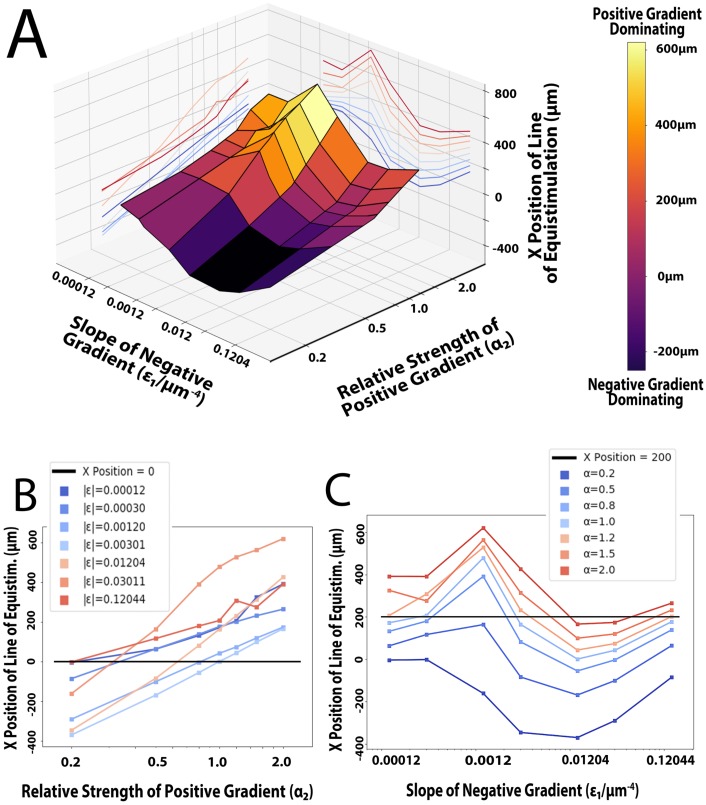
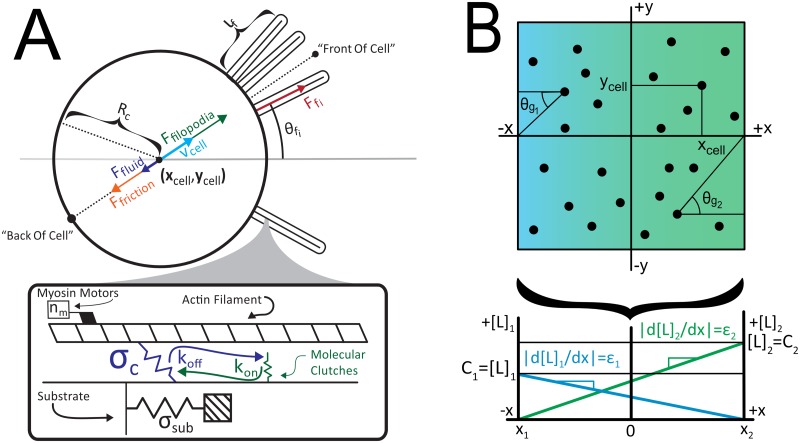
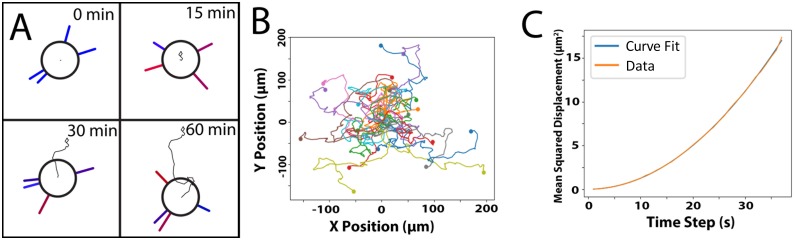
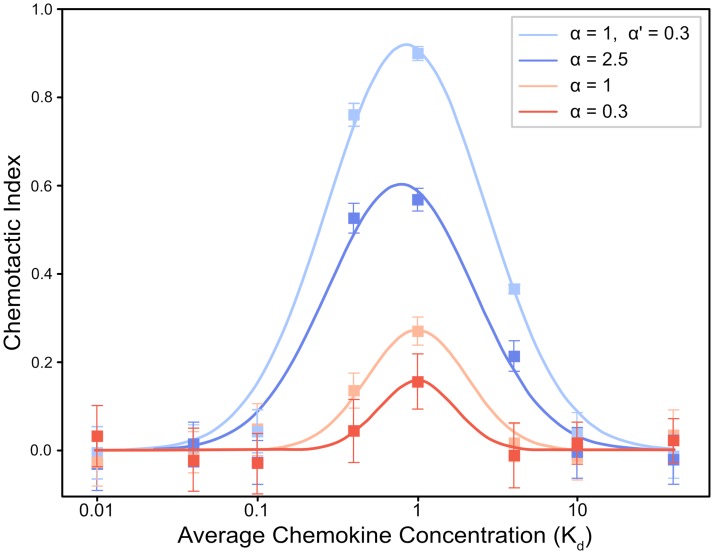
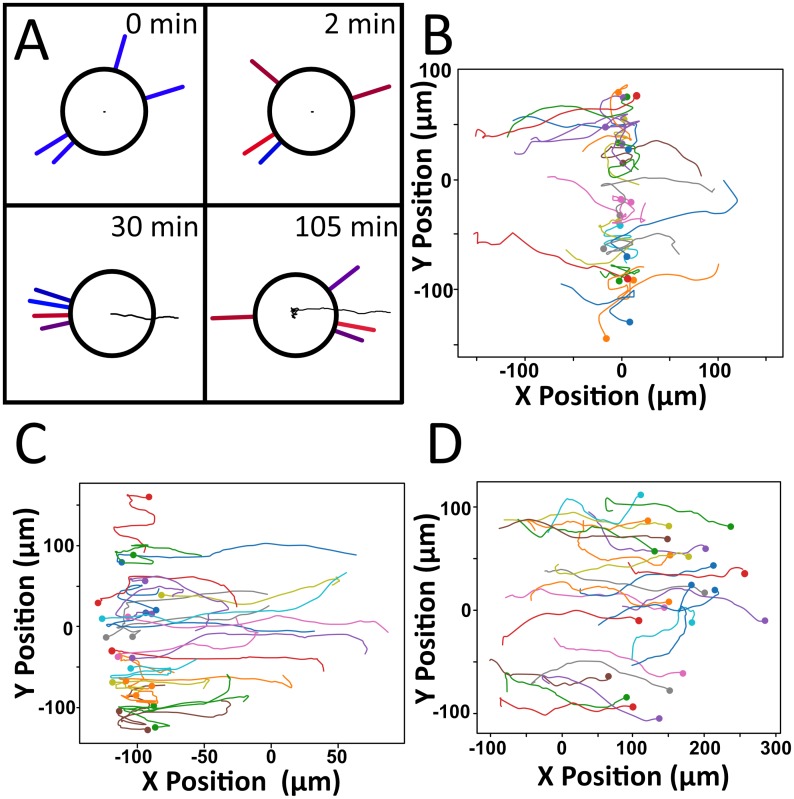
Dendritic cells (DCs) are the most effective professional antigen-presenting cell. They ferry antigen from the extremities to T cells and are essential for the initiation of an adaptive immune response. Despite interest in how DCs respond to chemical stimuli, there have been few attempts to model DC migration. In this paper, we simulate the motility of DCs by modeling the generation of forces by filopodia and a force balance on the cell. The direction of fliopodial extension is coupled to differential occupancy of cognate chemokine receptors across the cell. Our model simulates chemokinesis and chemotaxis in a variety of chemical and mechanical environments. Simulated DCs undergoing chemokinesis were measured to have a speed of 5.1 ± 0.07 μm·min-1 and a persistence time of 3.2 ± 0.46 min, consistent with experiment. Cells undergoing chemotaxis exhibited a stronger chemotactic response when exposed to lower average chemokine concentrations, also consistent with experiment. We predicted that when placed in two opposing gradients, cells will cluster in a line, which we call the "line of equistimulation;" this clustering has also been observed. We calculated the effect of varying gradient steepness on the line of equistimulation, with steeper gradients resulting in tighter clustering. Moreover, gradients are found to be most potent when cells are in a gradient of chemokine whose mean concentration is close to the binding of the Kd to the receptor, and least potent when the mean concentration is 0.1Kd. Comparing our simulations to experiment, we can give a quantitative measure of the strength of certain chemokines relative to others. Assigning the signal of CCL19 binding CCR7 a baseline strength of 1, we found CCL21 binding CCR7 had a strength of 0.28, and CXCL12 binding CXCR4 had a strength of 0.30. These differences emerge despite both chemokines having virtually the same Kd, suggesting a mechanism of signal amplification in DCs requiring further study.
树突状细胞 (DCs) 是最有效的专业抗原呈递细胞。它们从末梢运送抗原到 T 细胞,并对启动适应性免疫反应至关重要。尽管人们对 DC 如何对化学刺激做出反应很感兴趣,但很少有尝试对 DC 迁移进行建模。在本文中,我们通过建模丝状伪足产生的力和细胞上的力平衡来模拟 DC 的运动。丝状伪足延伸的方向与细胞上同源趋化因子受体的差异占据相关联。我们的模型模拟了各种化学和机械环境中的趋化运动和趋化运动。模拟的 DC 进行趋化运动的速度为 5.1 ± 0.07 μm·min-1,持续时间为 3.2 ± 0.46 min,与实验结果一致。进行趋化运动的细胞在暴露于较低平均趋化浓度时表现出更强的趋化反应,这也与实验结果一致。我们预测,当放置在两个相反的梯度中时,细胞将聚集在一条线上,我们称之为“等刺激线”;这种聚集也已经被观察到。我们计算了梯度陡度对等刺激线的影响,梯度陡度越大,聚集越紧密。此外,当细胞处于接近 Kd 与受体结合的趋化浓度的梯度中时,梯度的作用最强,而当平均浓度为 0.1Kd 时,梯度的作用最弱。将我们的模拟与实验进行比较,我们可以对某些趋化因子相对于其他趋化因子的强度进行定量测量。将 CCL19 结合 CCR7 的信号指定为基本强度 1,我们发现 CCL21 结合 CCR7 的强度为 0.28,而 CXCL12 结合 CXCR4 的强度为 0.30。尽管这两种趋化因子的 Kd 几乎相同,但仍出现了这些差异,这表明 DC 中存在信号放大机制,需要进一步研究。