Department of Chemistry and Biochemistry, Baylor University, Waco, United States.
Elife. 2019 Oct 29;8:e46096. doi: 10.7554/eLife.46096.
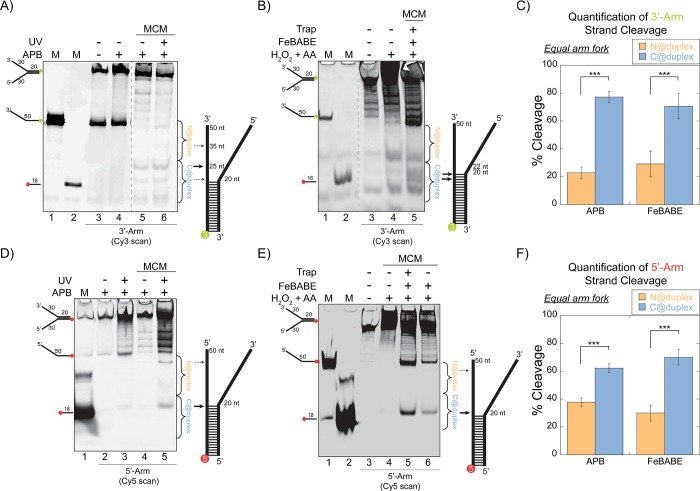
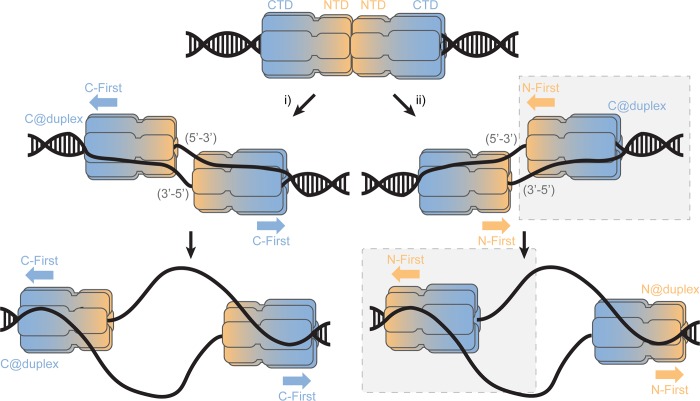
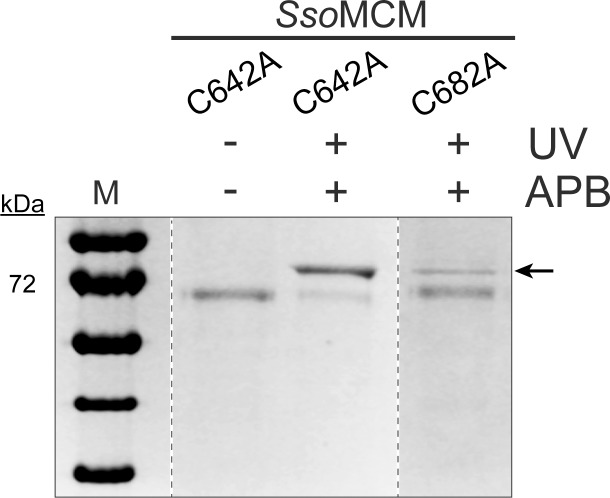
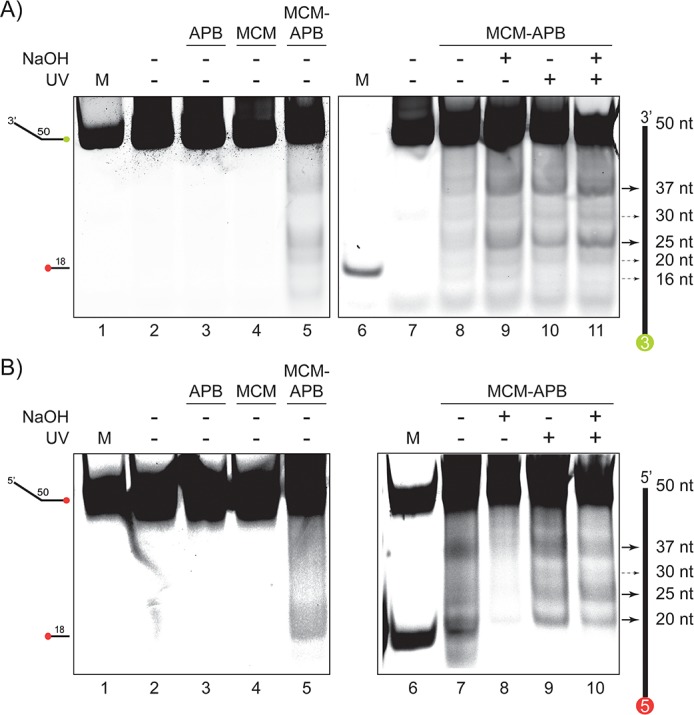
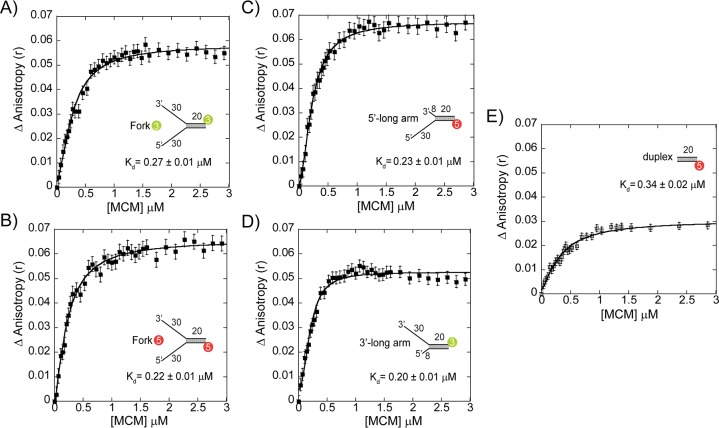
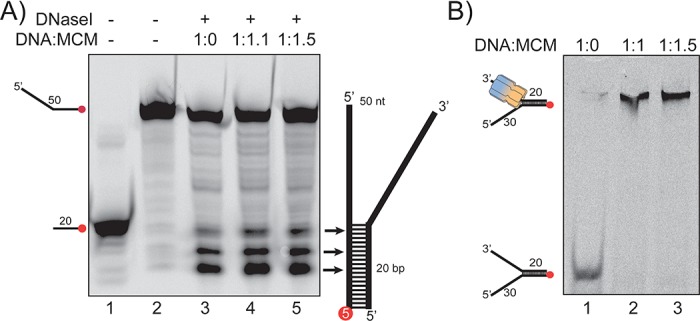
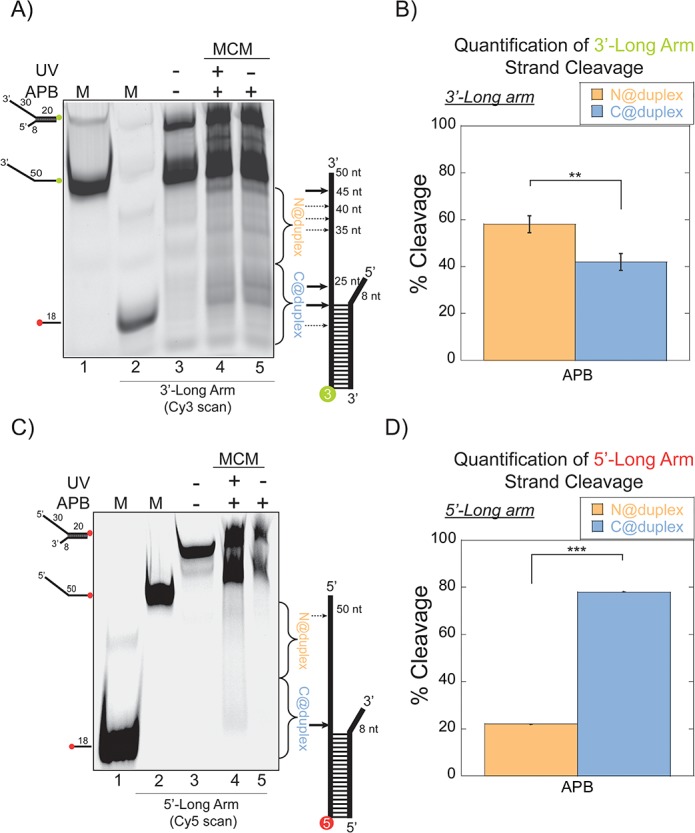
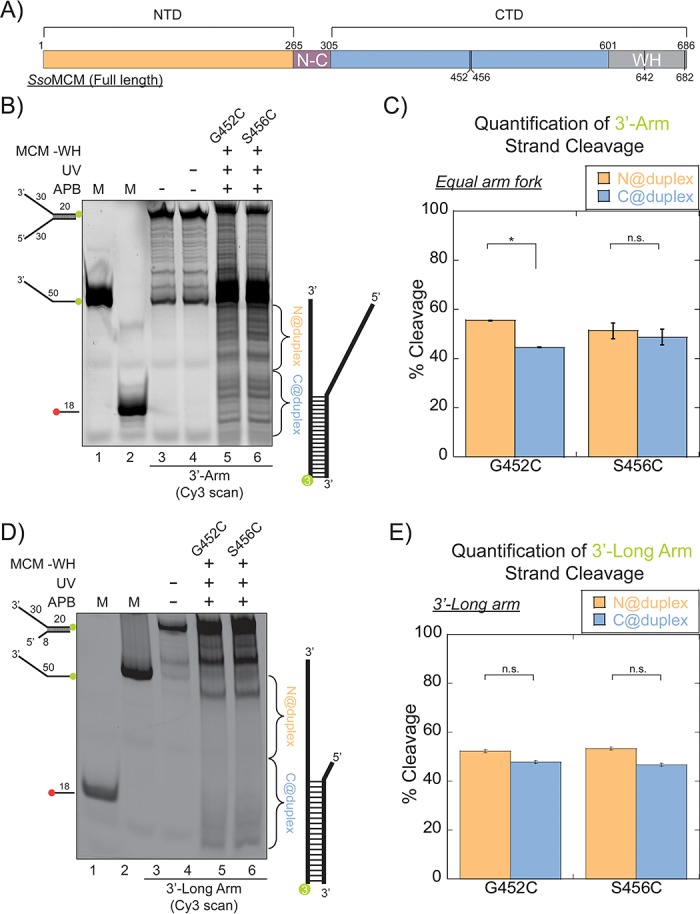
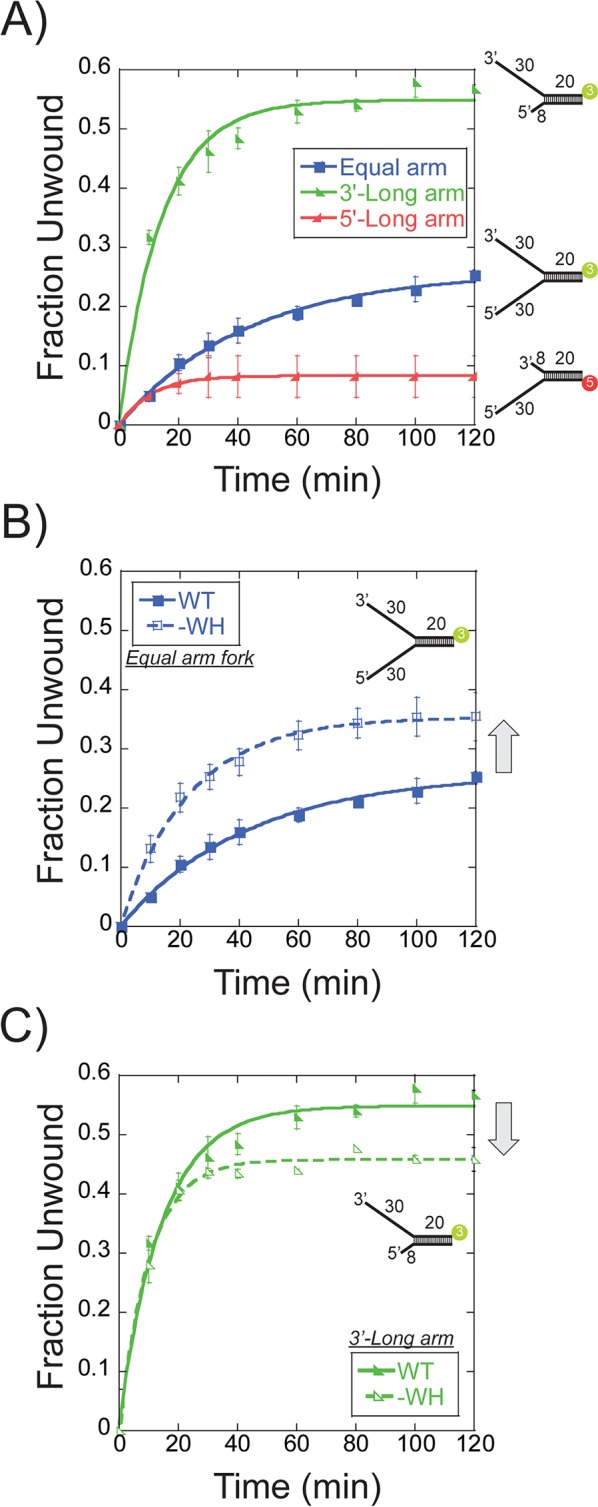
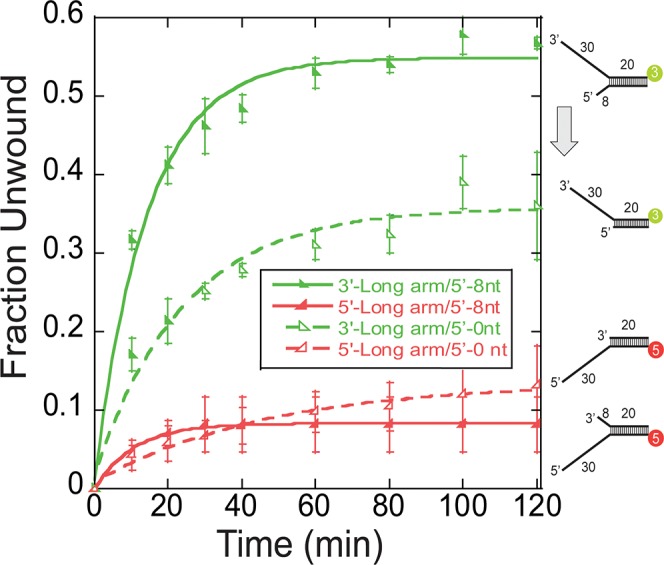
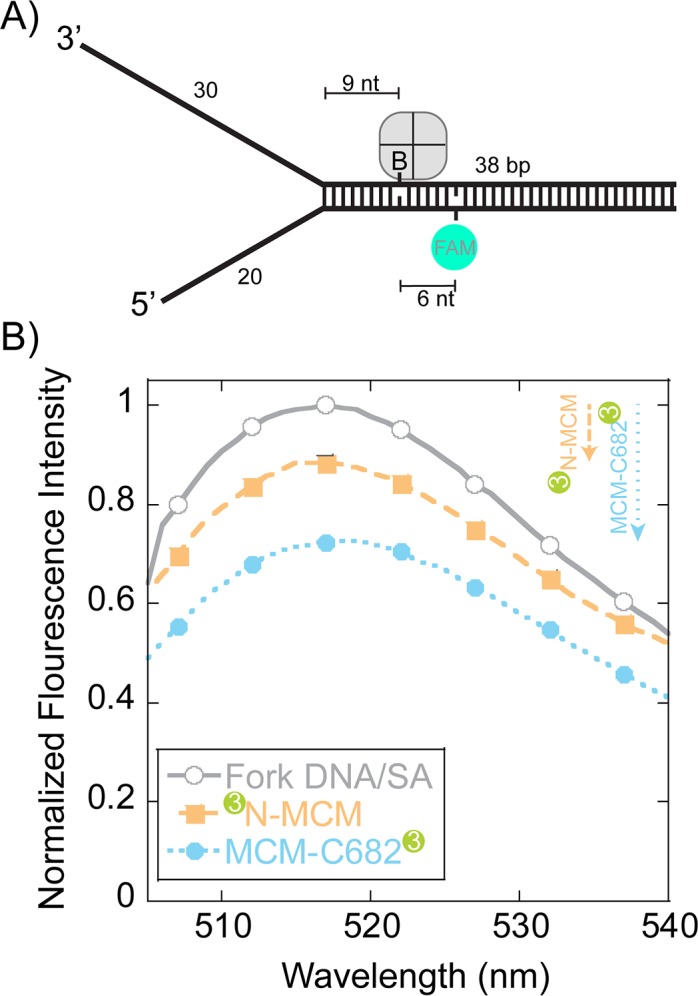
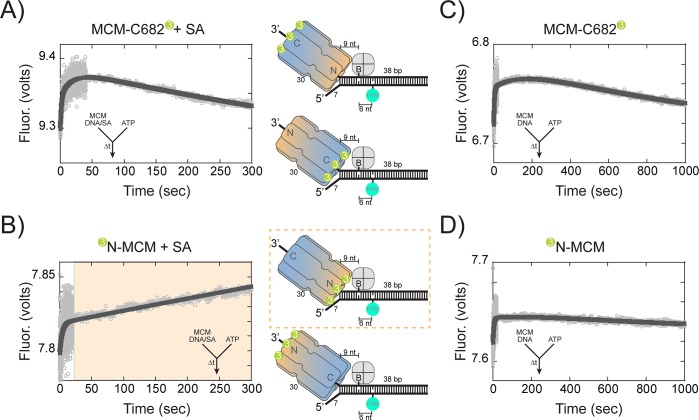
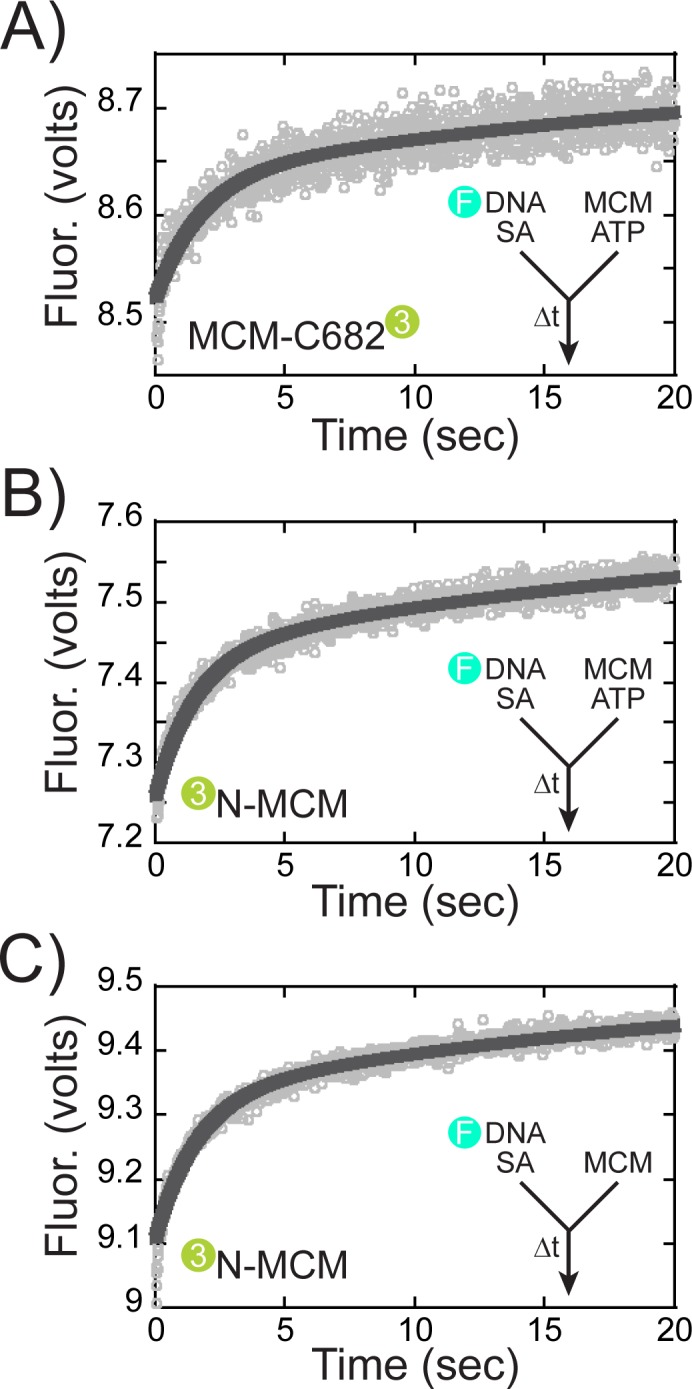
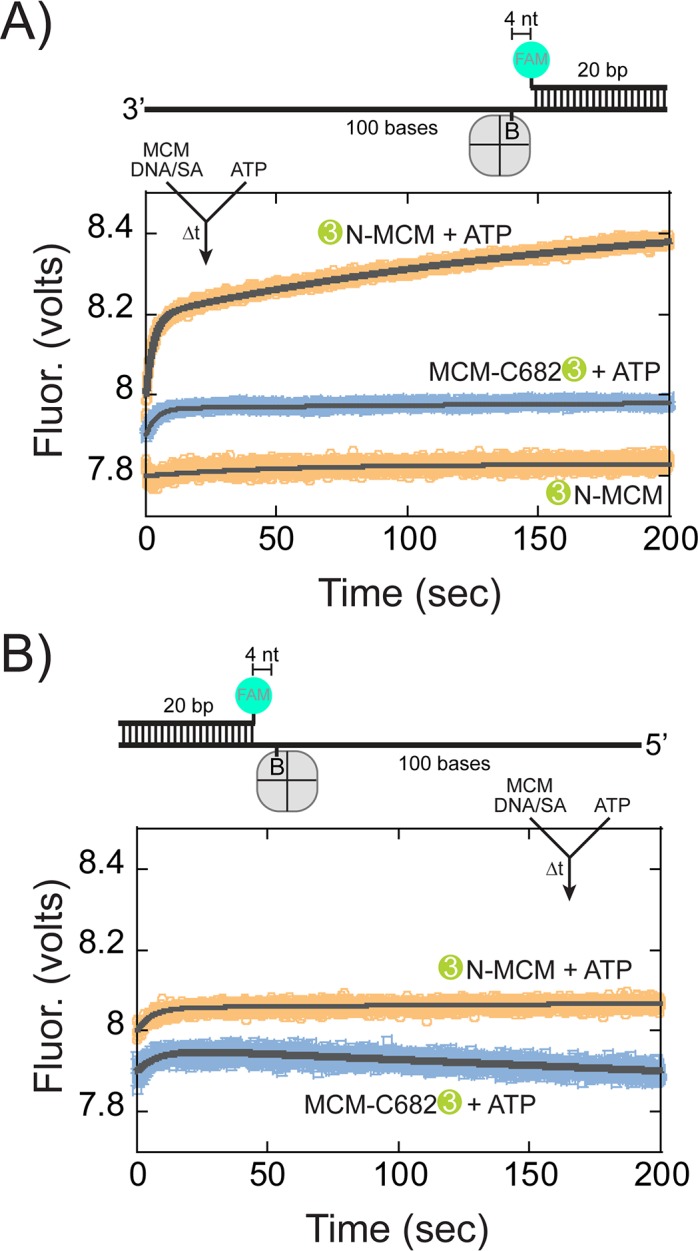
DNA replication requires that the duplex genomic DNA strands be separated; a function that is implemented by ring-shaped hexameric helicases in all Domains. Helicases are composed of two domains, an N- terminal DNA binding domain (NTD) and a C- terminal motor domain (CTD). Replication is controlled by loading of helicases at origins of replication, activation to preferentially encircle one strand, and then translocation to begin separation of the two strands. Using a combination of site-specific DNA footprinting, single-turnover unwinding assays, and unique fluorescence translocation monitoring, we have been able to quantify the binding distribution and the translocation orientation of (formally ) MCM on DNA. Our results show that both the DNA substrate and the C-terminal winged-helix (WH) domain influence the orientation but that translocation on DNA proceeds N-first.
DNA 复制需要将双链基因组 DNA 链分离;这一功能由所有领域的环形六聚体解旋酶来执行。解旋酶由两个结构域组成,一个 N 端 DNA 结合结构域(NTD)和一个 C 端马达结构域(CTD)。复制通过在复制起点处加载解旋酶来控制,优先激活其中一条链,然后进行易位以开始分离两条链。我们使用特定于位点的 DNA 足迹法、单轮解旋测定和独特的荧光易位监测相结合的方法,能够定量(形式上)MCM 在 DNA 上的结合分布和易位取向。我们的结果表明,DNA 底物和 C 端翼状螺旋(WH)结构域都影响取向,但 DNA 上的易位是 N 端优先进行的。