Department of Medicine, University of California San Diego, San Diego, United States.
The Second Affiliated Hospital of Guangzhou Medical University (GMU), The State Key Laboratory of Respiratory Disease, Guangdong Provincial Key Laboratory of Allergy & Clinical Immunology, Guangzhou, China.
Elife. 2020 Feb 4;9:e49416. doi: 10.7554/eLife.49416.
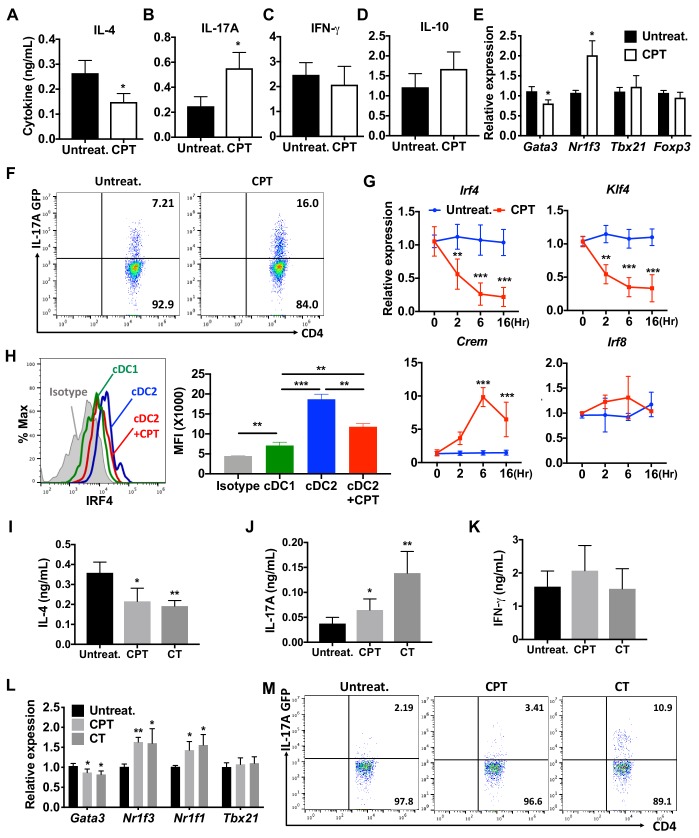
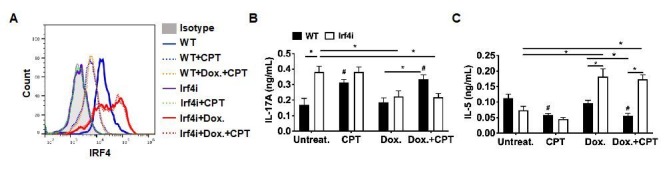
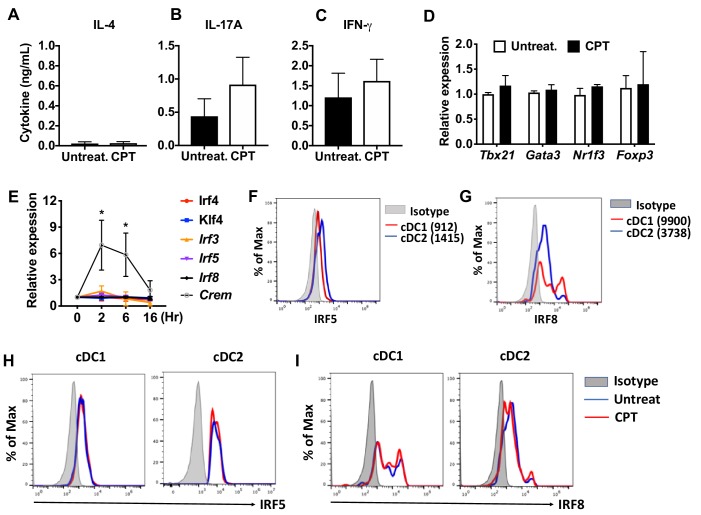
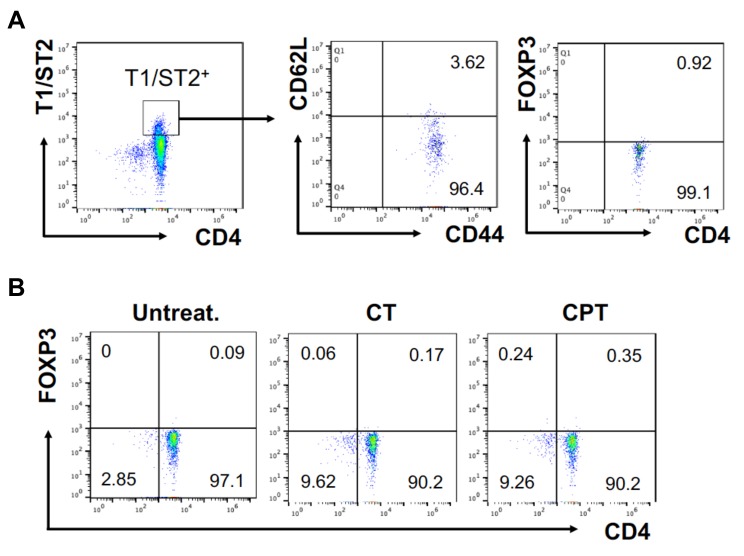
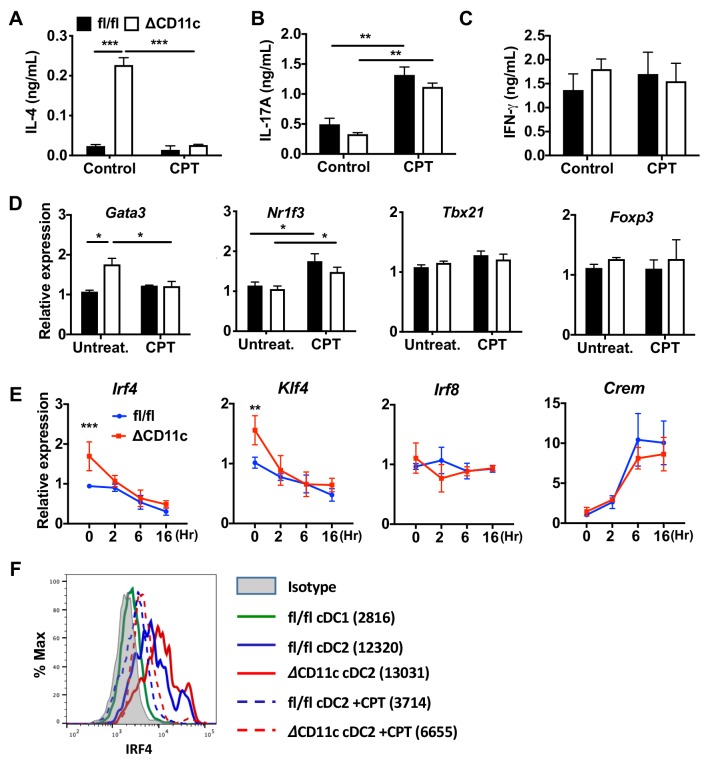
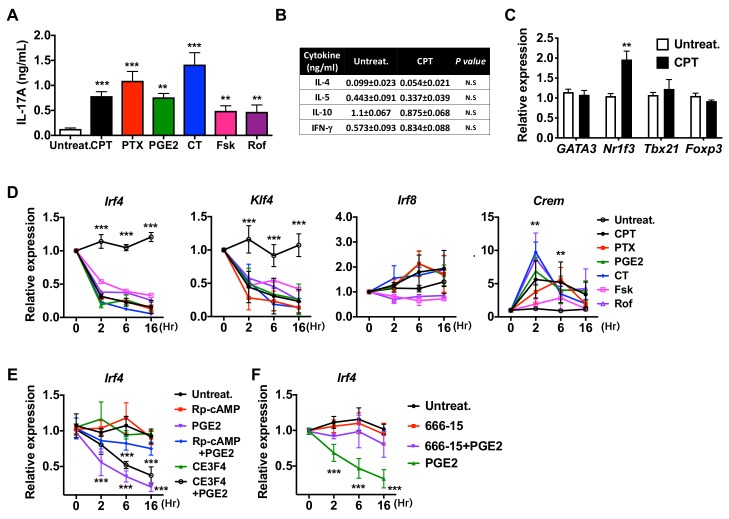
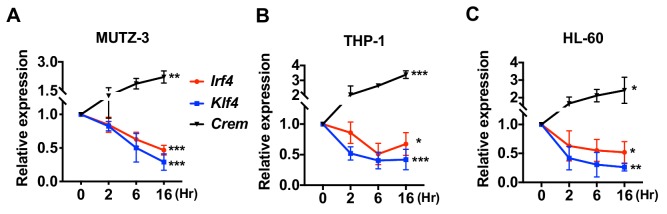
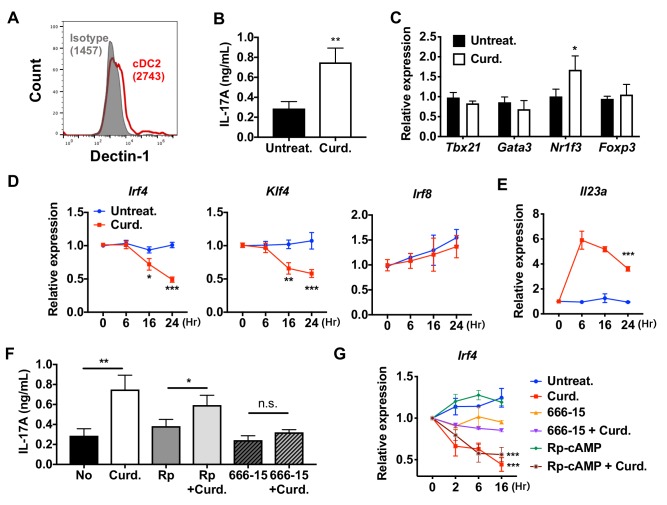
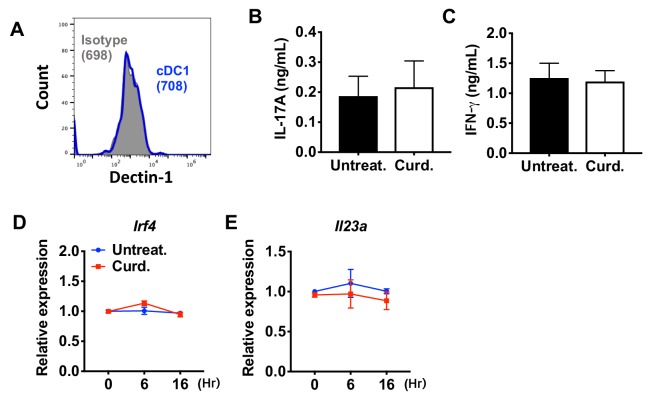
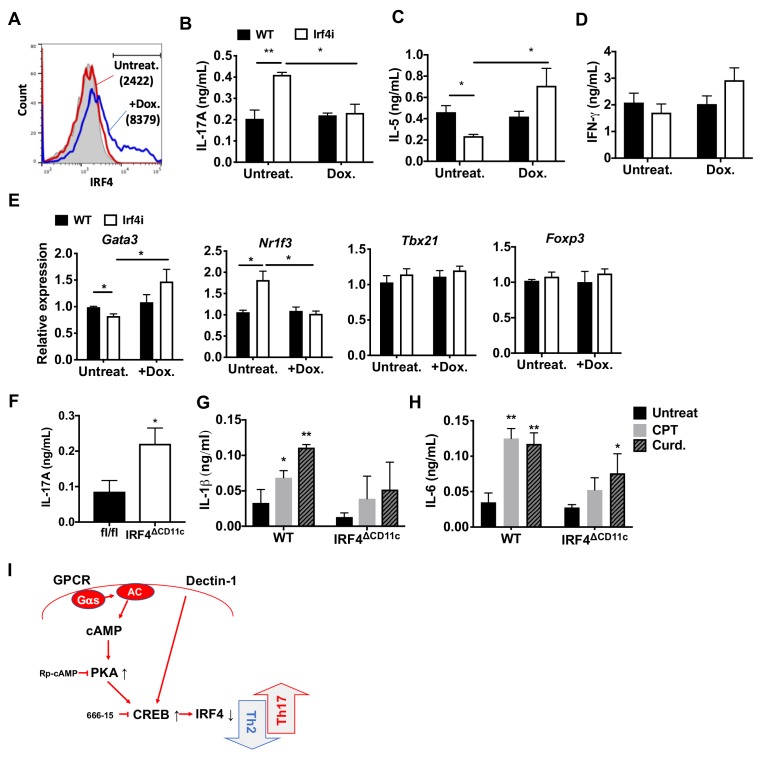
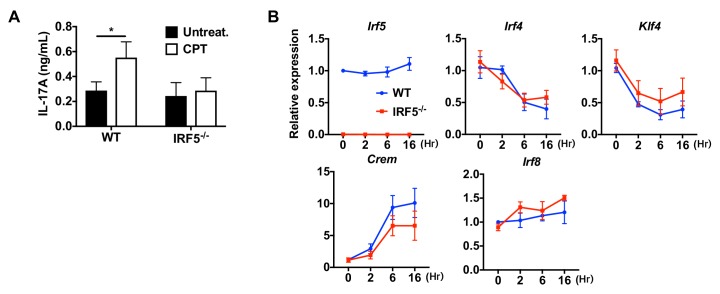
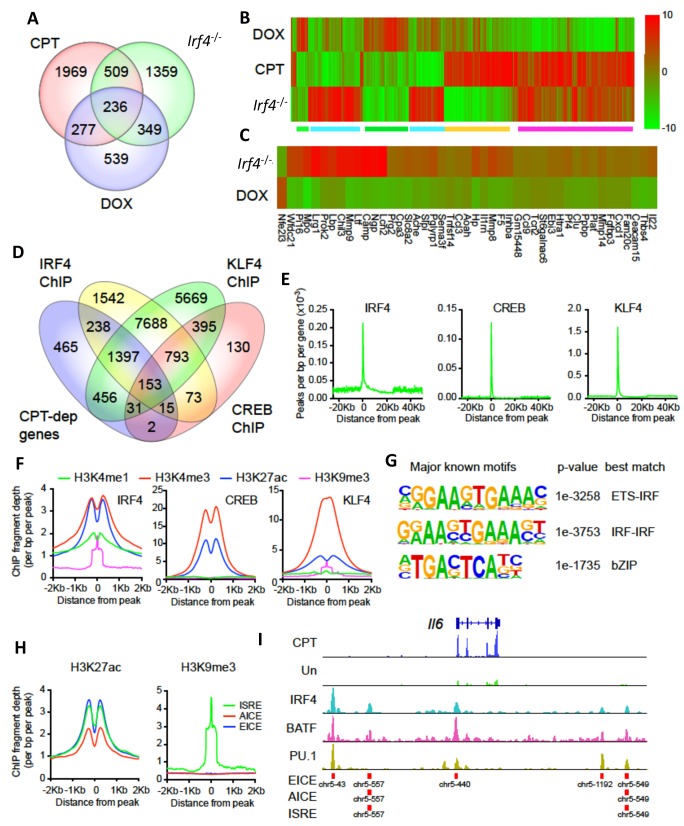
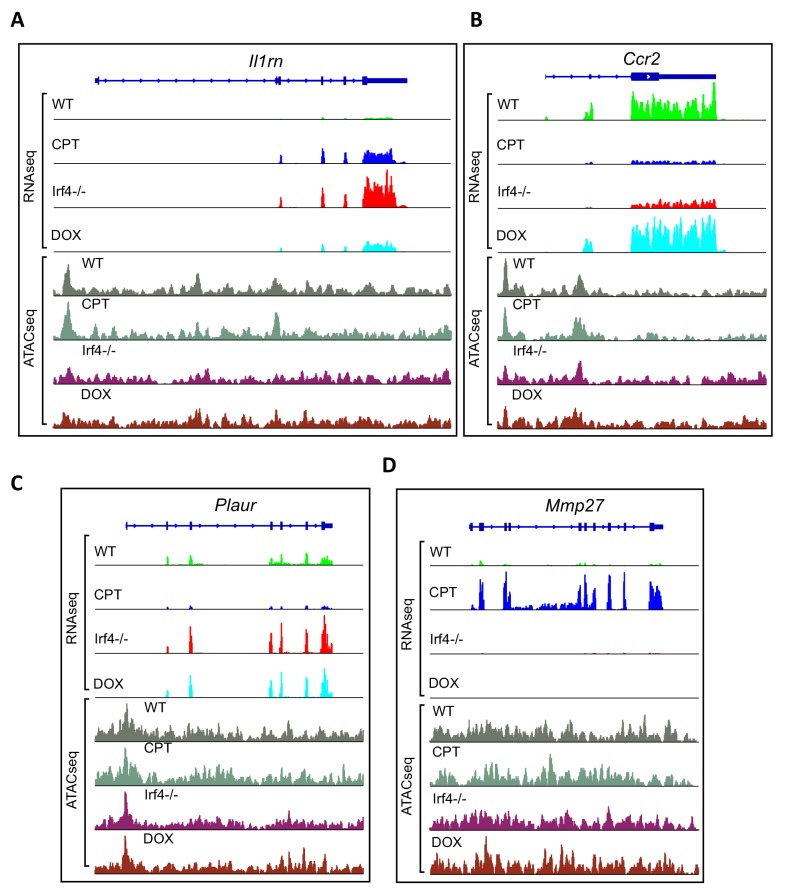
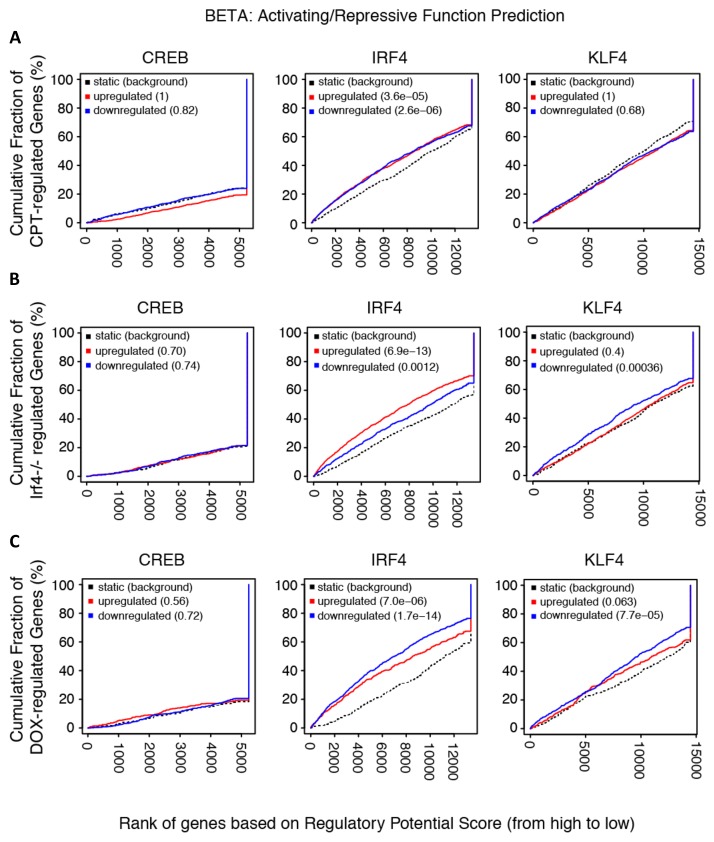
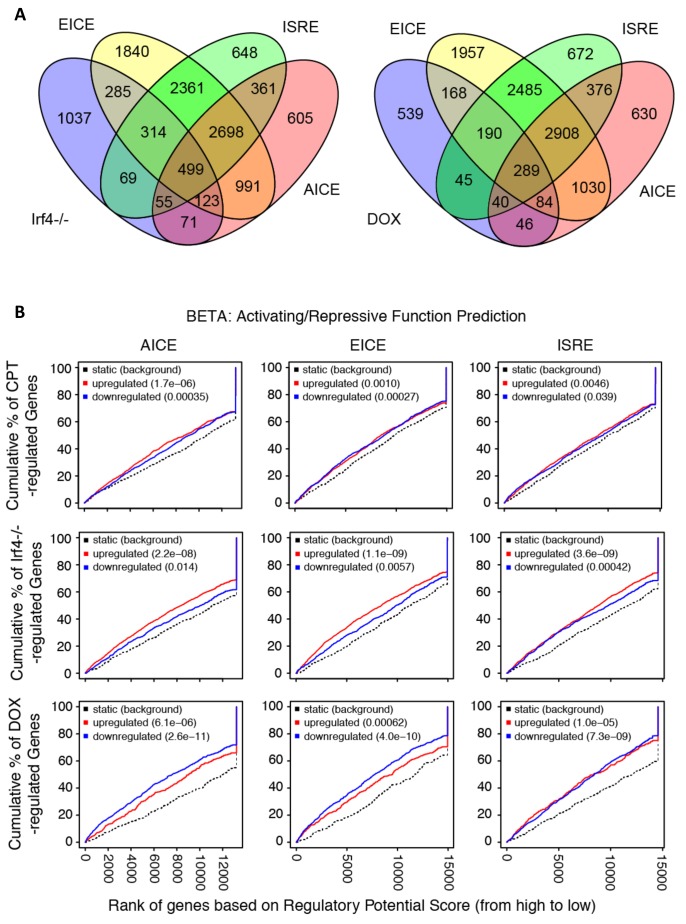
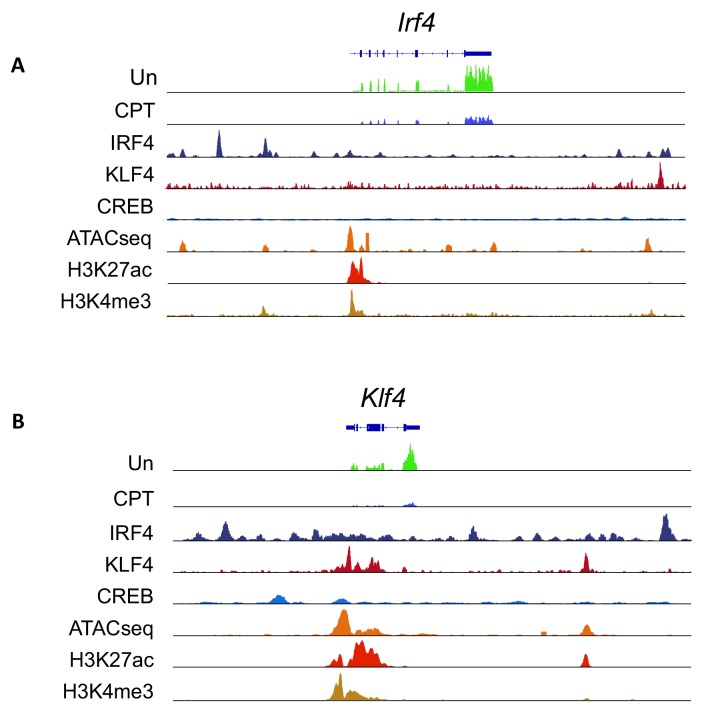
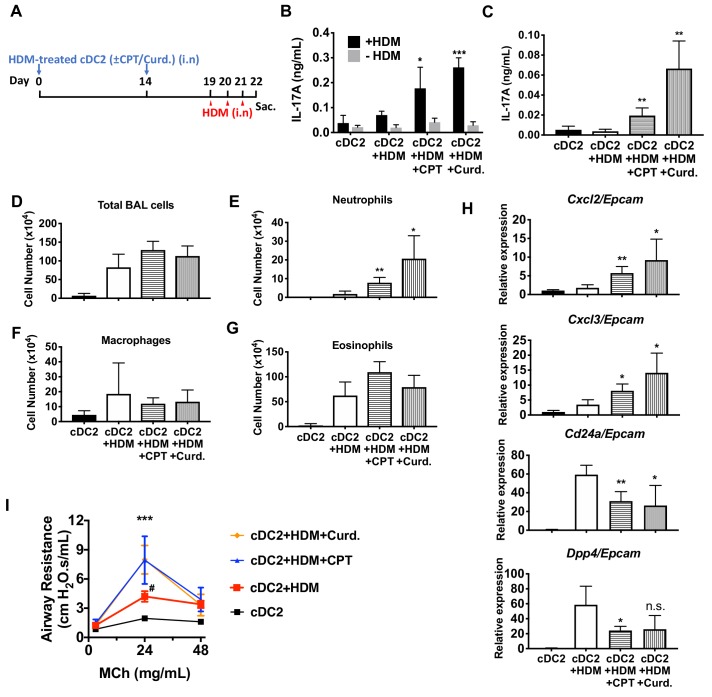
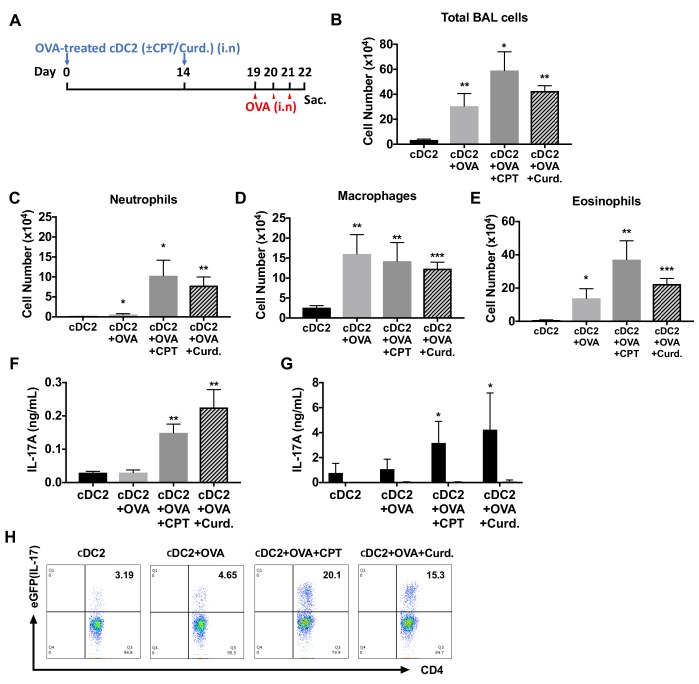
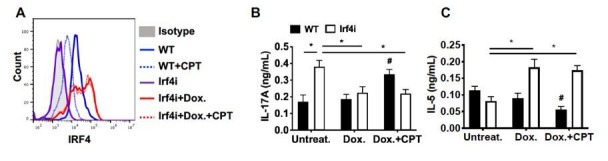
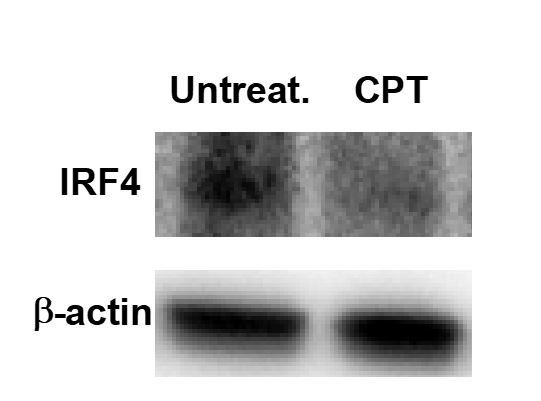
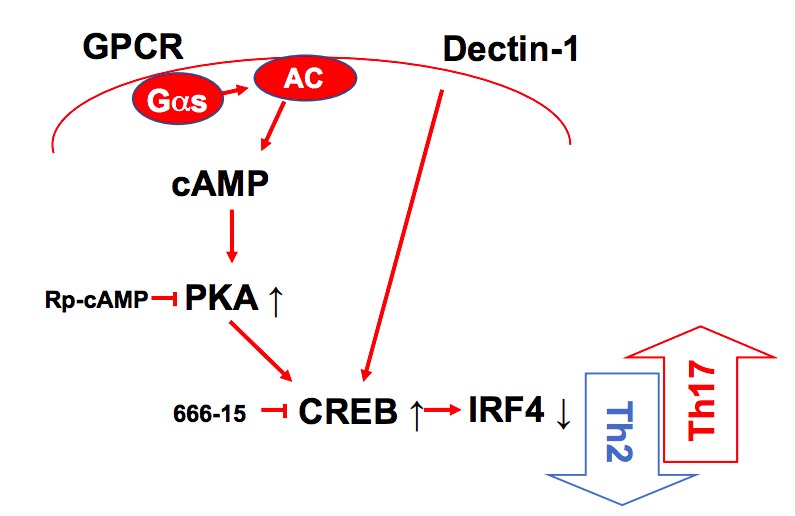
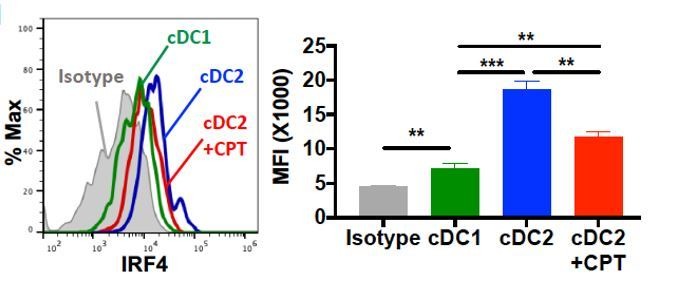
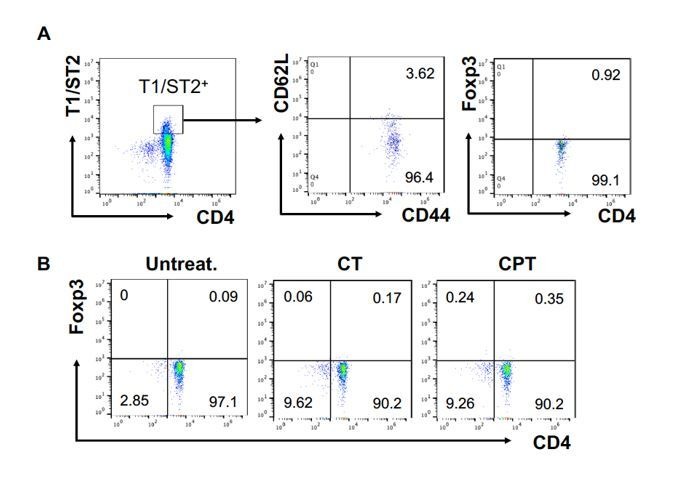
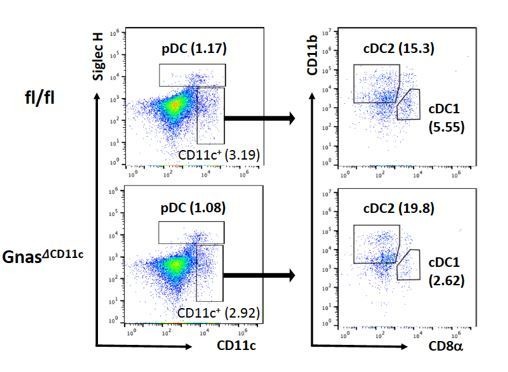
Cyclic AMP (cAMP) is involved in many biological processes but little is known regarding its role in shaping immunity. Here we show that cAMP-PKA-CREB signaling (a pattern recognition receptor [PRR]-independent mechanism) regulates conventional type-2 Dendritic Cells (cDC2s) in mice and reprograms their Th17-inducing properties via repression of IRF4 and KLF4, transcription factors essential for cDC2-mediated Th2 induction. In mice, genetic loss of IRF4 phenocopies the effects of cAMP on Th17 induction and restoration of IRF4 prevents the cAMP effect. Moreover, curdlan, a PRR-dependent microbial product, activates CREB and represses IRF4 and KLF4, resulting in a pro-Th17 phenotype of cDC2s. These in vitro and in vivo results define a novel signaling pathway by which cDC2s display plasticity and provide a new molecular basis for the classification of novel cDC2 and cDC17 subsets. The findings also reveal that repressing IRF4 and KLF4 pathway can be harnessed for immuno-regulation.
环磷酸腺苷 (cAMP) 参与许多生物过程,但对于其在塑造免疫中的作用知之甚少。在这里,我们表明 cAMP-PKA-CREB 信号(一种模式识别受体 [PRR]-非依赖性机制)调节小鼠中的常规 2 型树突状细胞 (cDC2),并通过抑制 IRF4 和 KLF4 来重新编程其 Th17 诱导特性,IRF4 和 KLF4 是 cDC2 介导的 Th2 诱导所必需的转录因子。在小鼠中,IRF4 的基因缺失可模拟 cAMP 对 Th17 诱导的影响,而 IRF4 的恢复可阻止 cAMP 的作用。此外,一种依赖于 PRR 的微生物产物,几丁质葡聚糖,可激活 CREB 并抑制 IRF4 和 KLF4,从而导致 cDC2 的 Th17 表型。这些体外和体内结果定义了一个新的信号通路,通过该通路,cDC2 显示出可塑性,并为新型 cDC2 和 cDC17 亚群的分类提供了新的分子基础。这些发现还表明,抑制 IRF4 和 KLF4 途径可用于免疫调节。