Burggren Warren W
Developmental Integrative Biology, Department of Biological Sciences, University of North Texas, Denton, TX, United States.
Front Physiol. 2020 Jan 22;10:1634. doi: 10.3389/fphys.2019.01634. eCollection 2019.
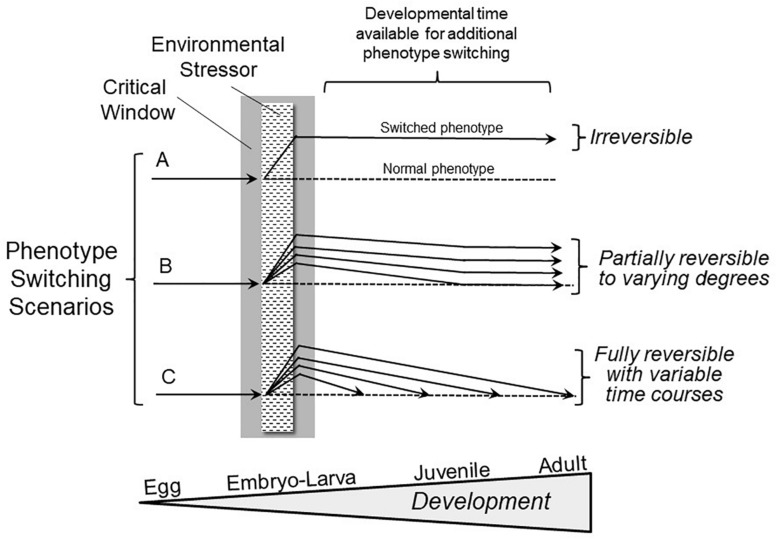
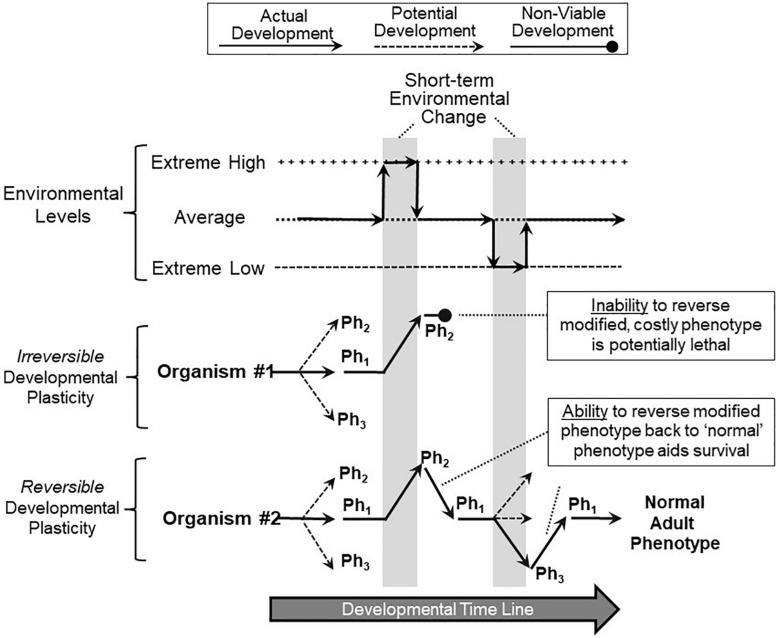
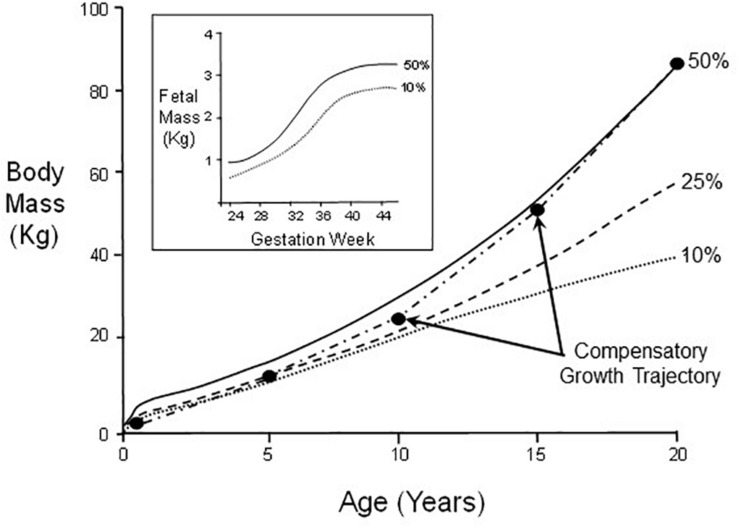
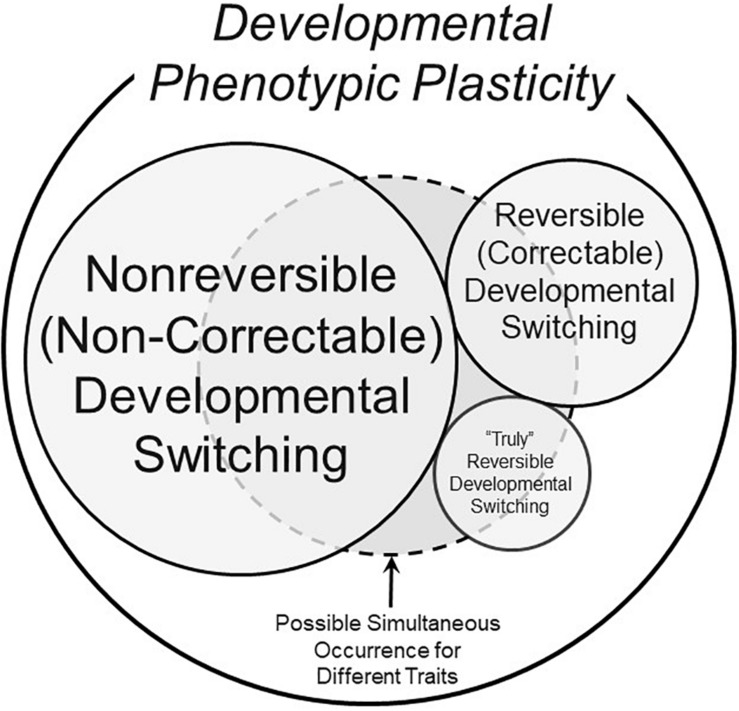
The prevalent view of developmental phenotypic switching holds that phenotype modifications occurring during critical windows of development are "irreversible" - that is, once produced by environmental perturbation, the consequent juvenile and/or adult phenotypes are indelibly modified. Certainly, many such changes appear to be non-reversible later in life. Yet, whether animals with switched phenotypes during early development are unable to return to a normal range of adult phenotypes, or whether they do not experience the specific environmental conditions necessary for them to switch back to the normal range of adult phenotypes, remains an open question. Moreover, developmental critical windows are typically brief, early periods punctuating a much longer period of overall development. This leaves open additional developmental time for reversal (correction) of a switched phenotype resulting from an adverse environment early in development. Such reversal could occur from right after the critical window "closes," all the way into adulthood. In fact, examples abound of the capacity to return to normal adult phenotypes following phenotypic changes enabled by earlier developmental plasticity. Such examples include cold tolerance in the fruit fly, developmental switching of mouth formation in a nematode, organization of the spinal cord of larval zebrafish, camouflage pigmentation formation in larval newts, respiratory chemosensitivity in frogs, temperature-metabolism relations in turtles, development of vascular smooth muscle and kidney tissue in mammals, hatching/birth weight in numerous vertebrates,. More extreme cases of actual reversal (not just correction) occur in invertebrates (e.g., hydrozoans, barnacles) that actually 'backtrack' along normal developmental trajectories from adults back to earlier developmental stages. While developmental phenotypic switching is often viewed as a permanent deviation from the normal range of developmental plans, the concept of developmental phenotypic switching should be expanded to include sufficient plasticity allowing subsequent correction resulting in the normal adult phenotype.
发育表型转换的普遍观点认为,在发育的关键窗口期发生的表型修饰是“不可逆的”——也就是说,一旦由环境扰动产生,随之而来的幼年和/或成年表型就会被永久性地改变。当然,许多这样的变化在生命后期似乎是不可逆的。然而,早期发育过程中表型发生转换的动物是无法恢复到正常的成年表型范围,还是它们没有经历恢复到正常成年表型范围所需的特定环境条件,仍然是一个悬而未决的问题。此外,发育关键窗口期通常很短暂,是贯穿整个更长发育时期的早期阶段。这就为逆转(纠正)发育早期由不利环境导致的转换表型留出了额外的发育时间。这种逆转可能在关键窗口“关闭”后立即发生,一直持续到成年期。事实上,有大量例子表明,早期发育可塑性导致表型变化后,动物有能力恢复到正常的成年表型。这些例子包括果蝇的耐寒性、线虫口器形成的发育转换、斑马鱼幼体脊髓的组织、蝾螈幼体的伪装色素形成、青蛙的呼吸化学敏感性、海龟的温度-代谢关系、哺乳动物血管平滑肌和肾脏组织的发育、众多脊椎动物的孵化/出生体重等。在无脊椎动物(如水螅、藤壶)中会出现更极端的实际逆转(不仅仅是纠正)情况,它们实际上会沿着正常发育轨迹从成体“回溯”到更早的发育阶段。虽然发育表型转换通常被视为对正常发育计划范围的永久性偏离,但发育表型转换的概念应扩展到包括足够的可塑性,允许随后的纠正,从而产生正常的成年表型。