Department of Biochemistry, University of Zurich, Zurich, Switzerland.
Institute of Biochemistry and Molecular Medicine, University of Bern, Bern, Switzerland.
Elife. 2020 May 6;9:e55853. doi: 10.7554/eLife.55853.
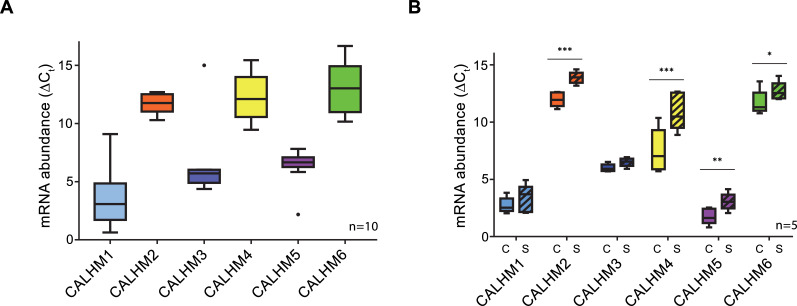
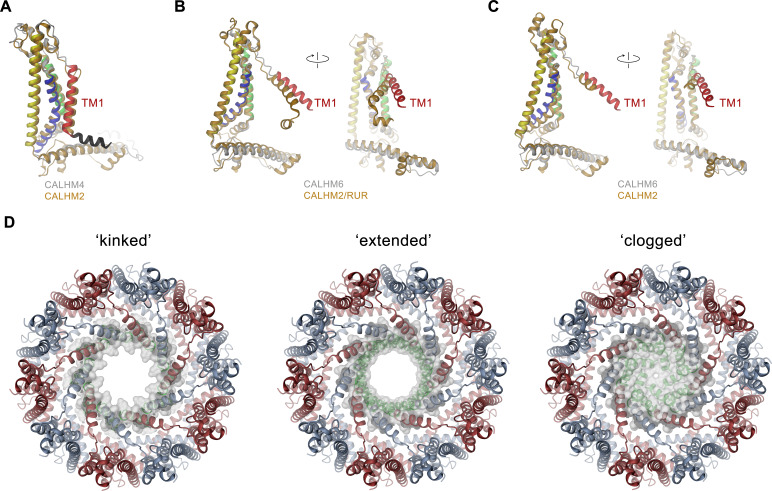
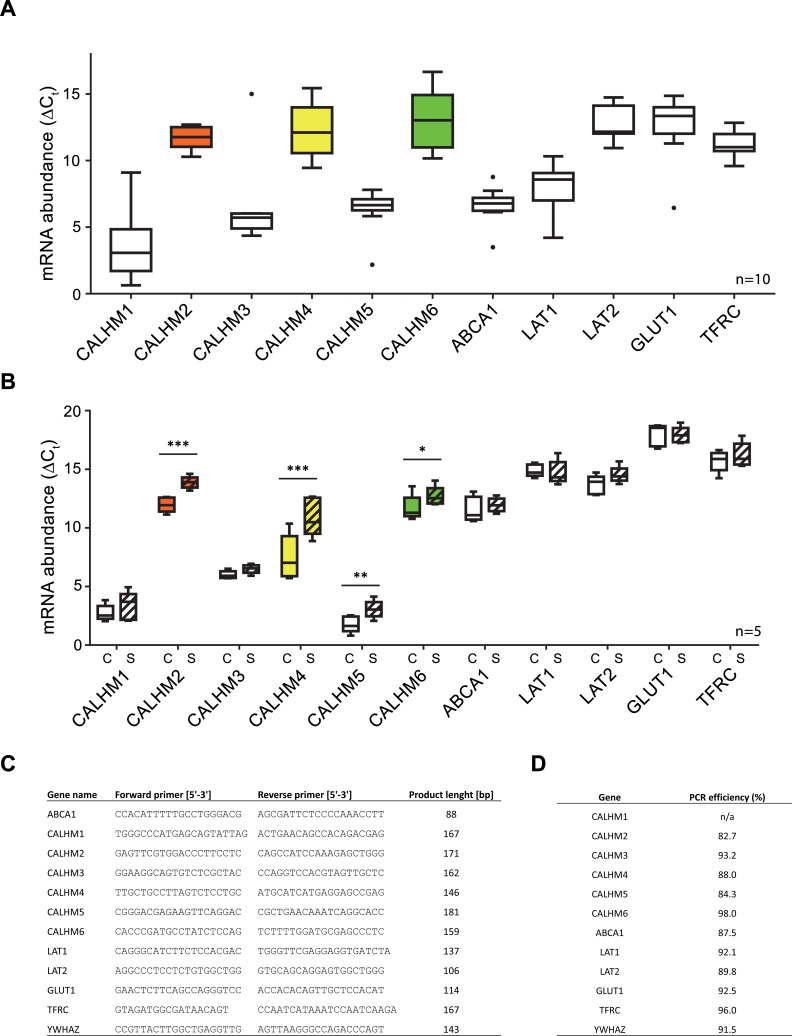
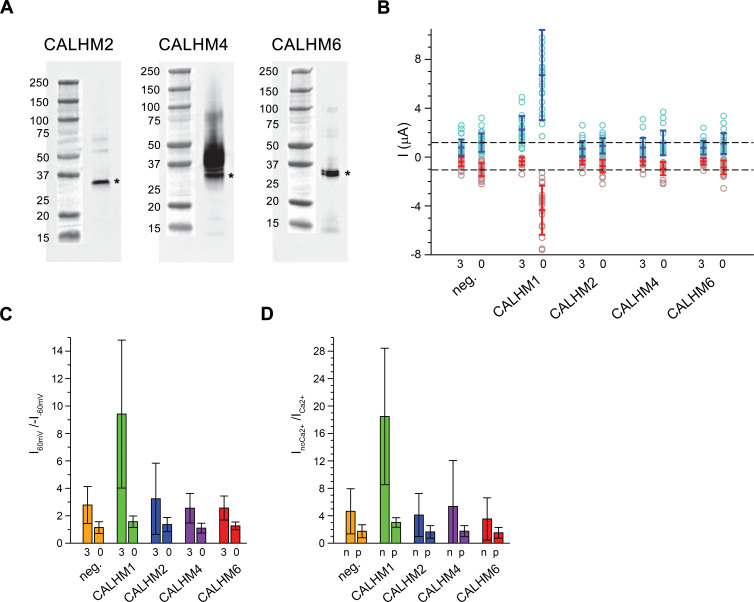
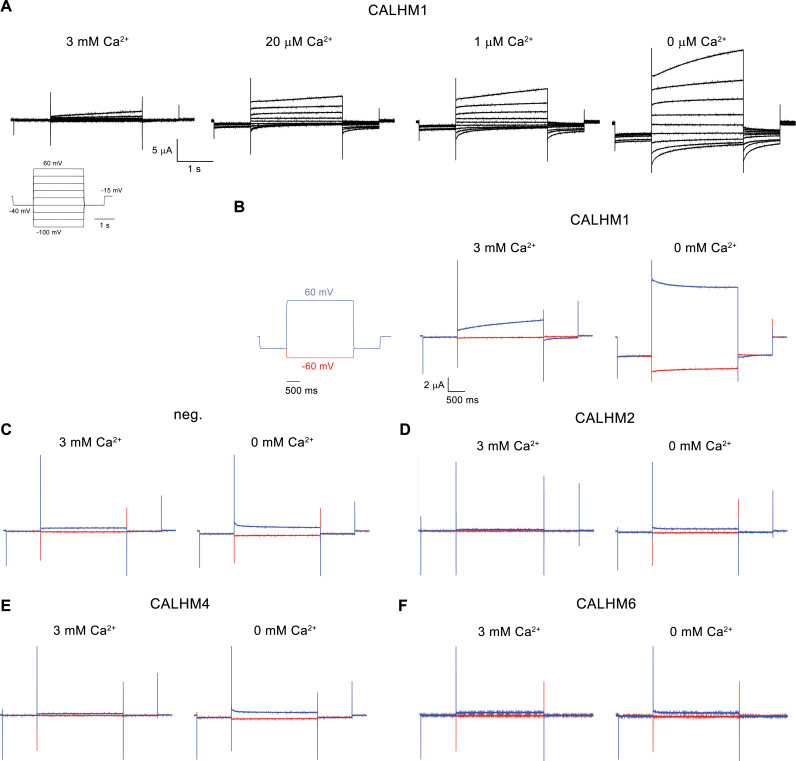
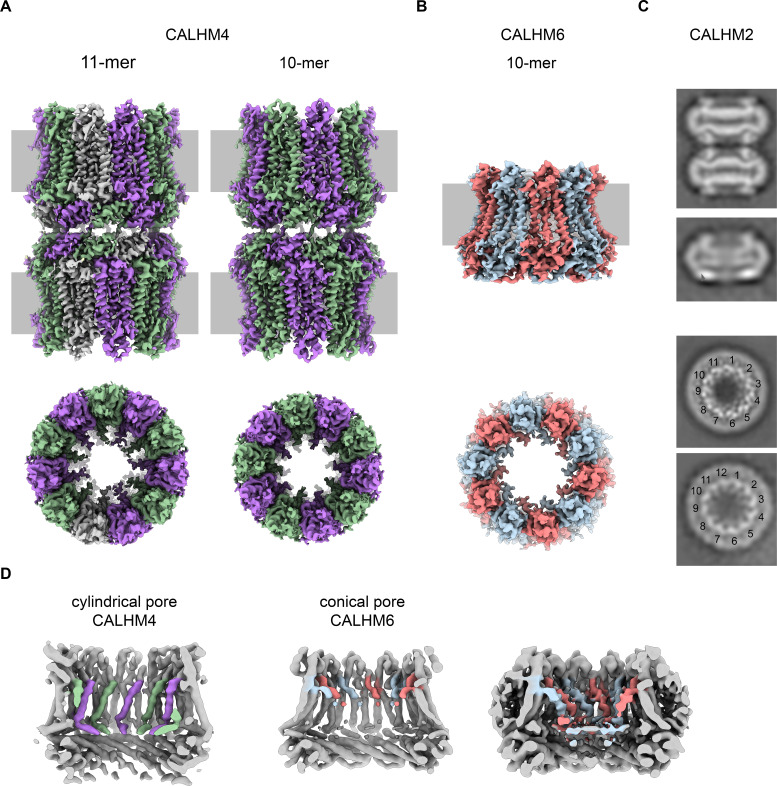
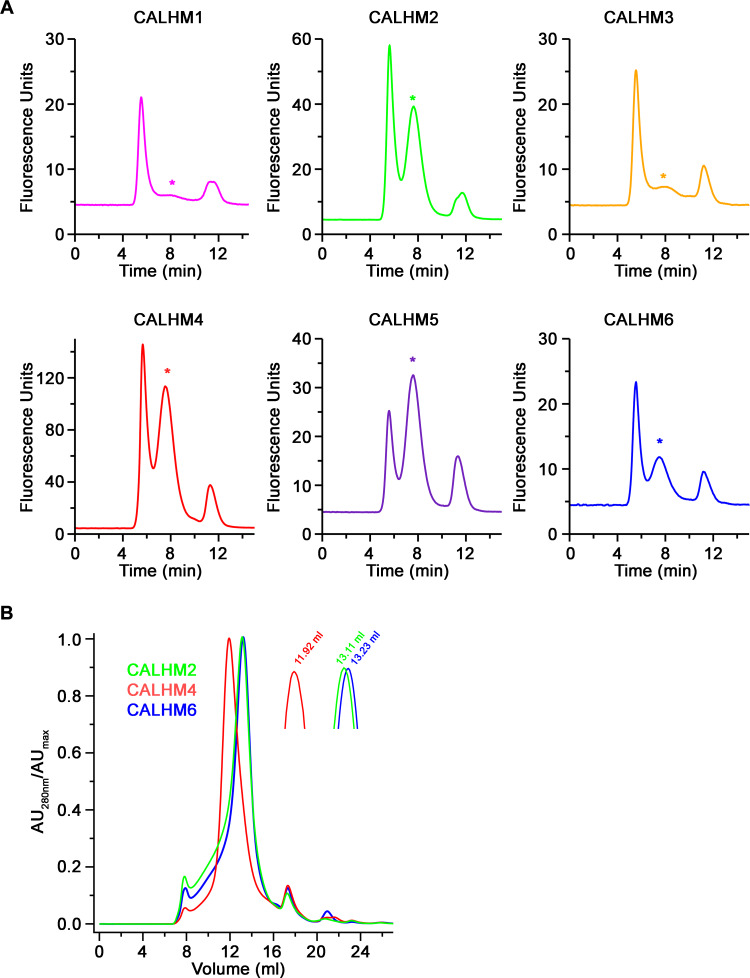
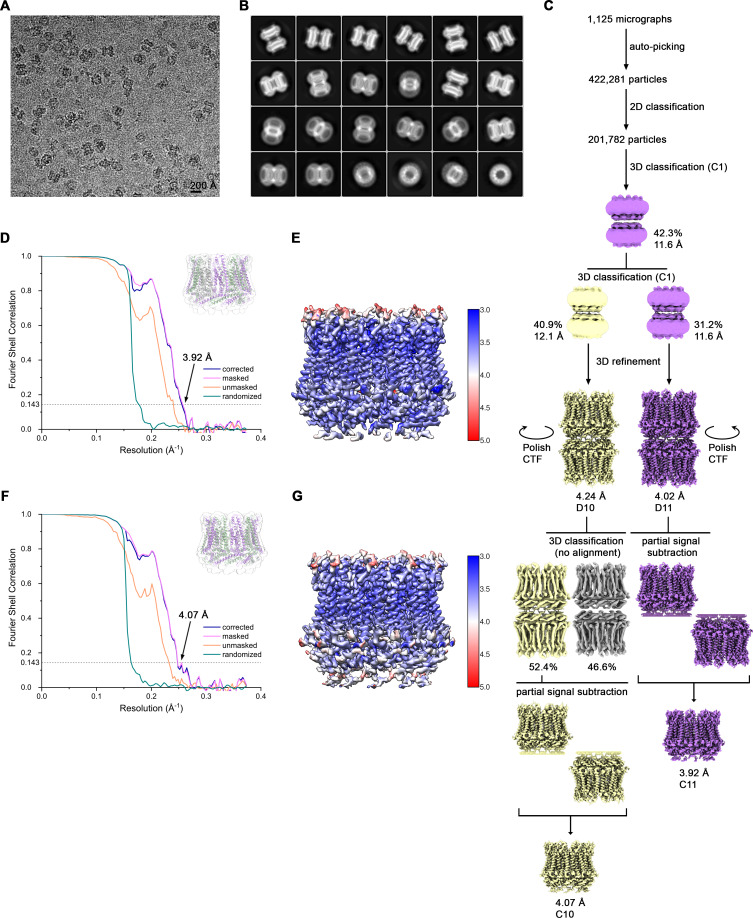
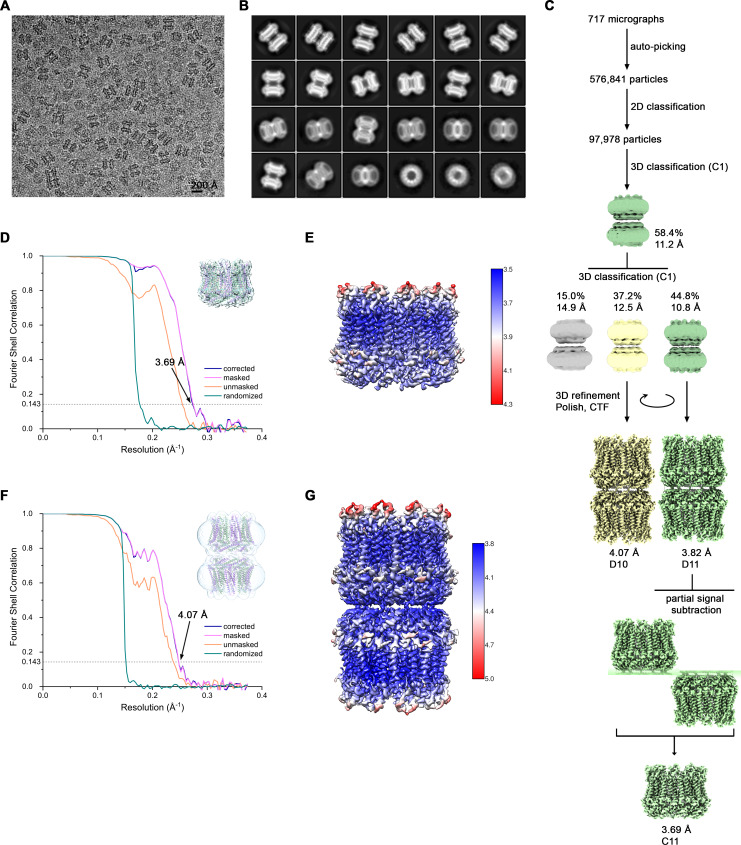
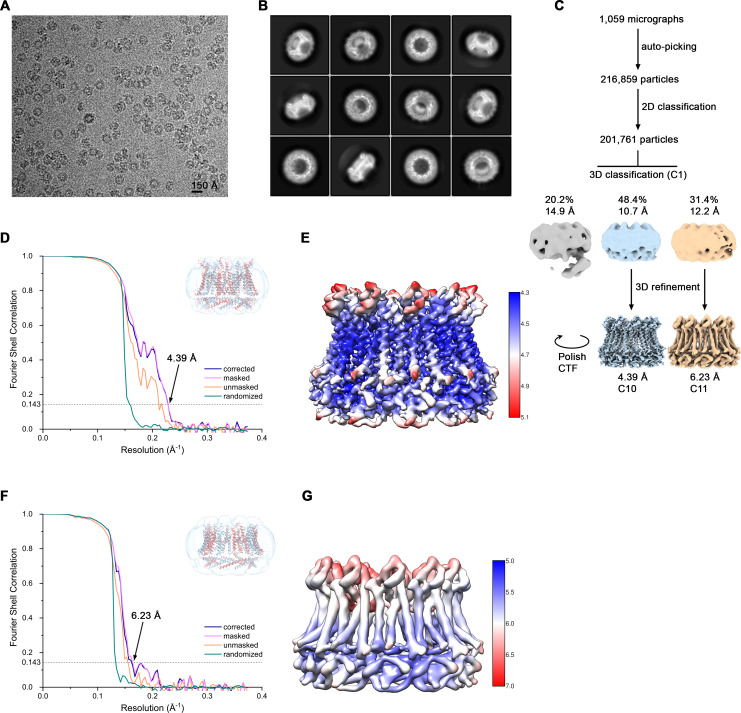
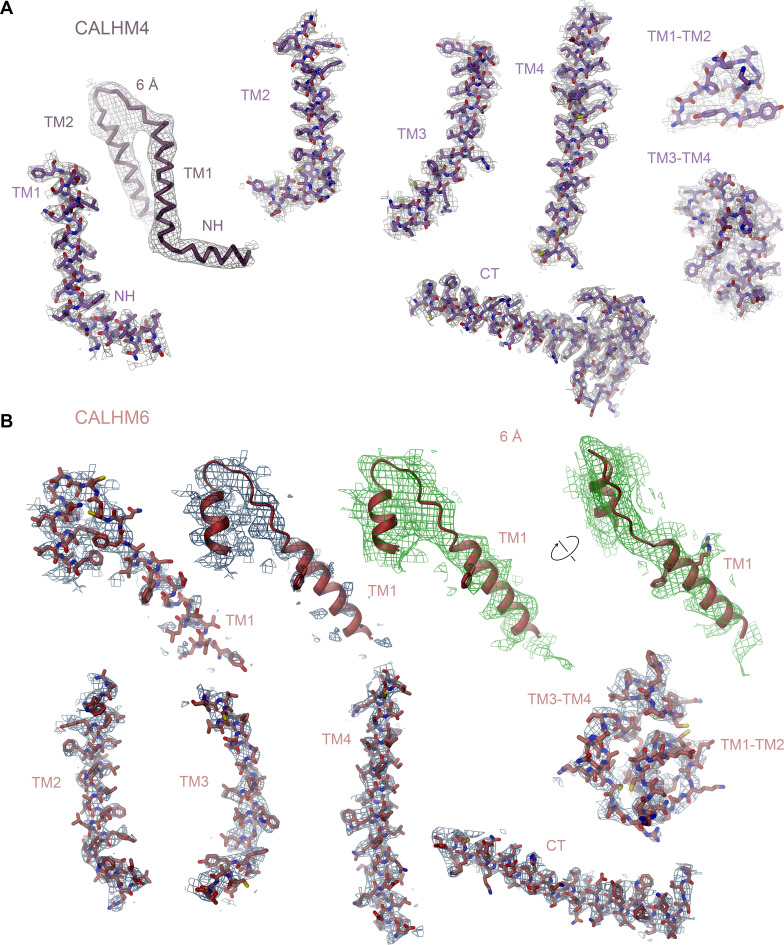
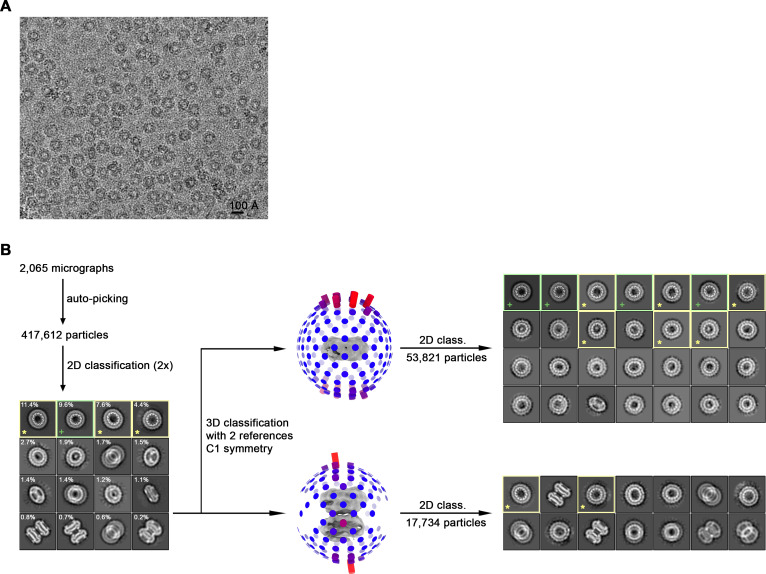
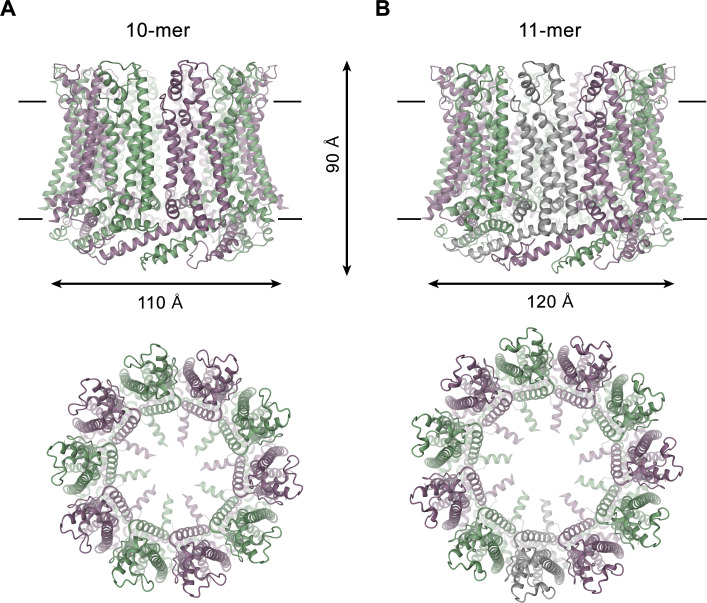
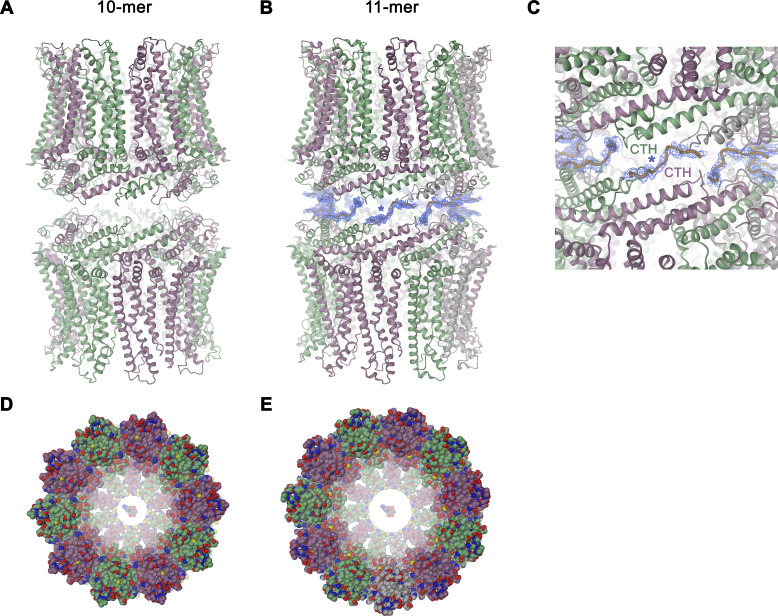
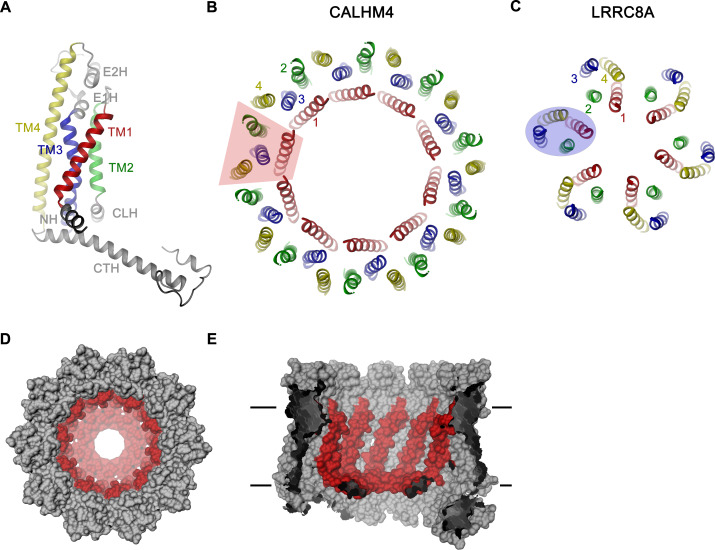
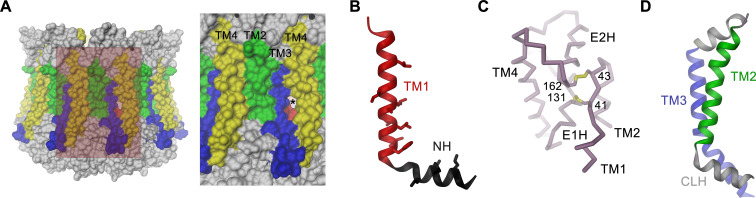
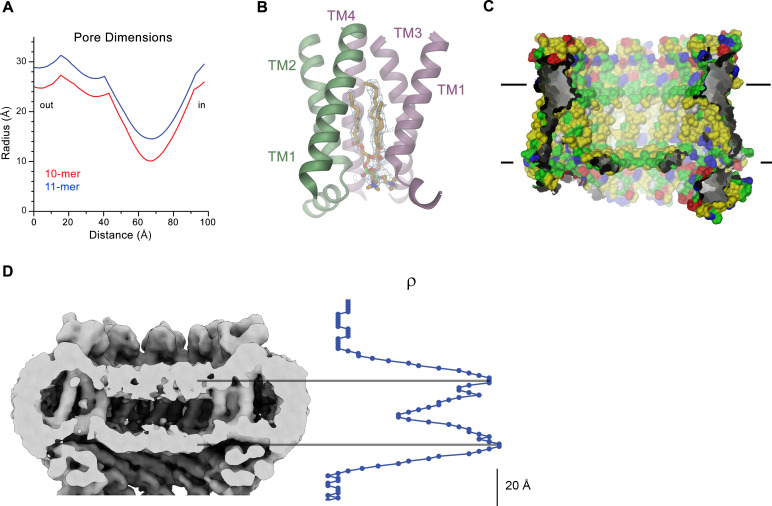
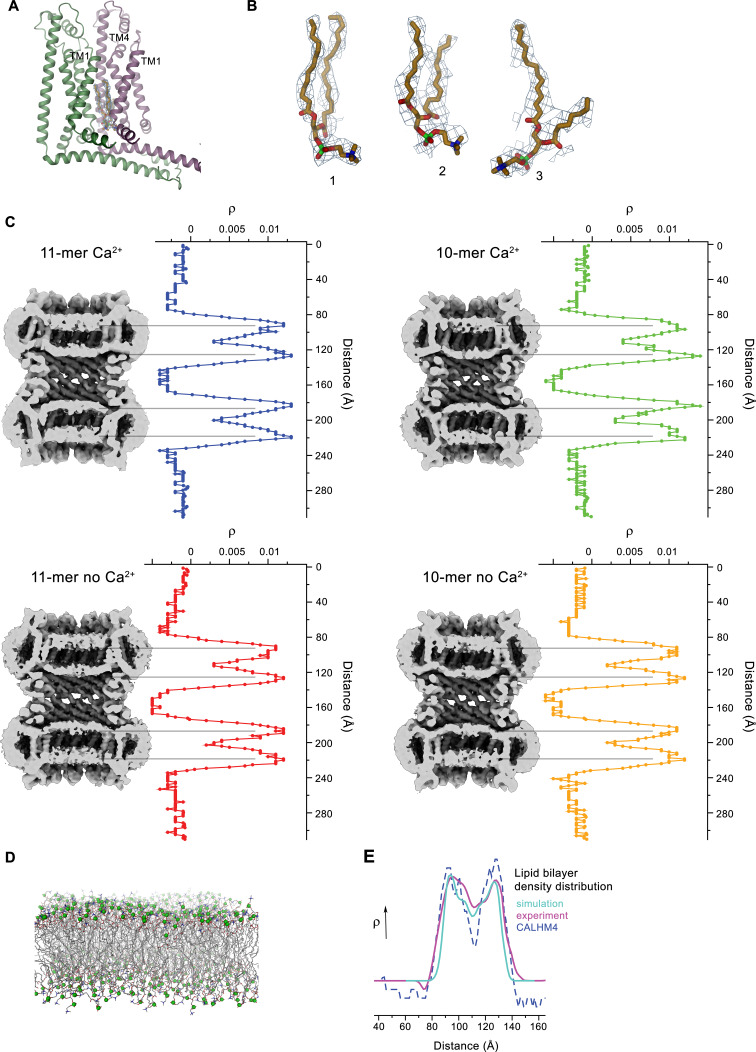
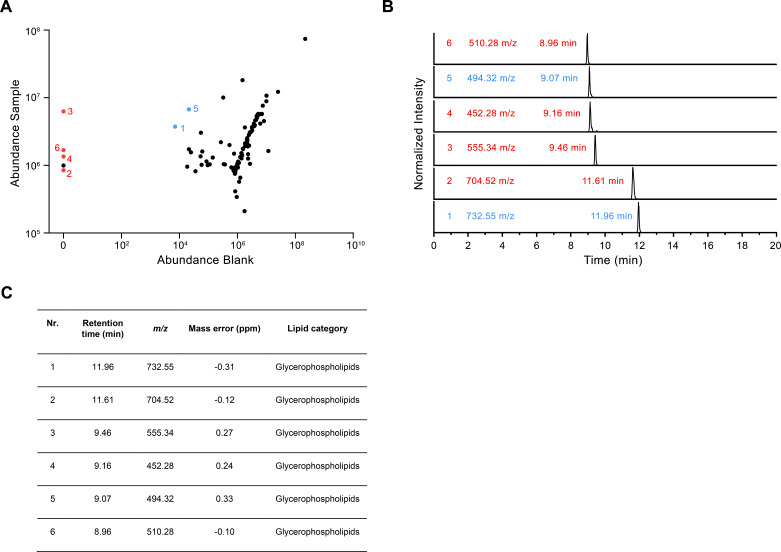
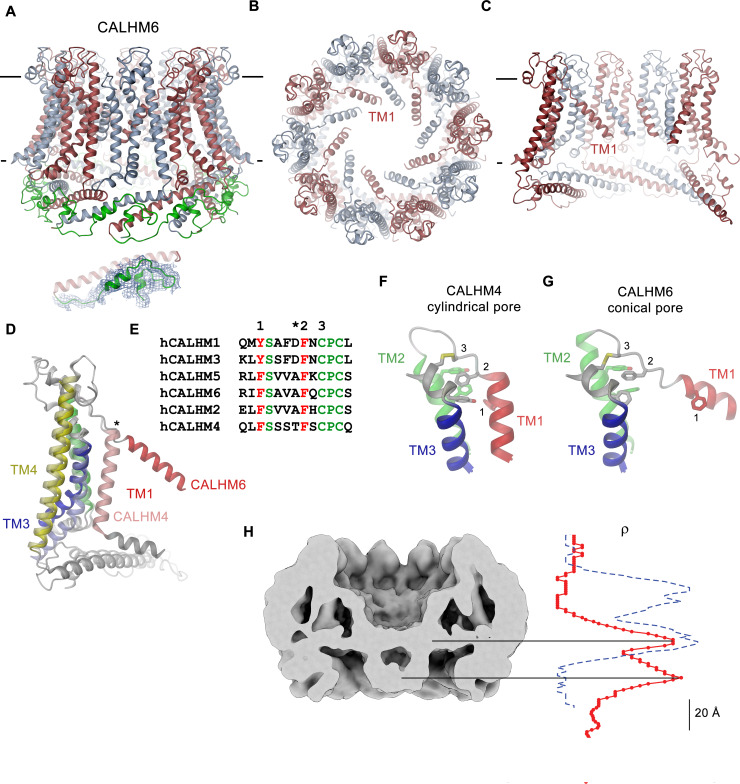
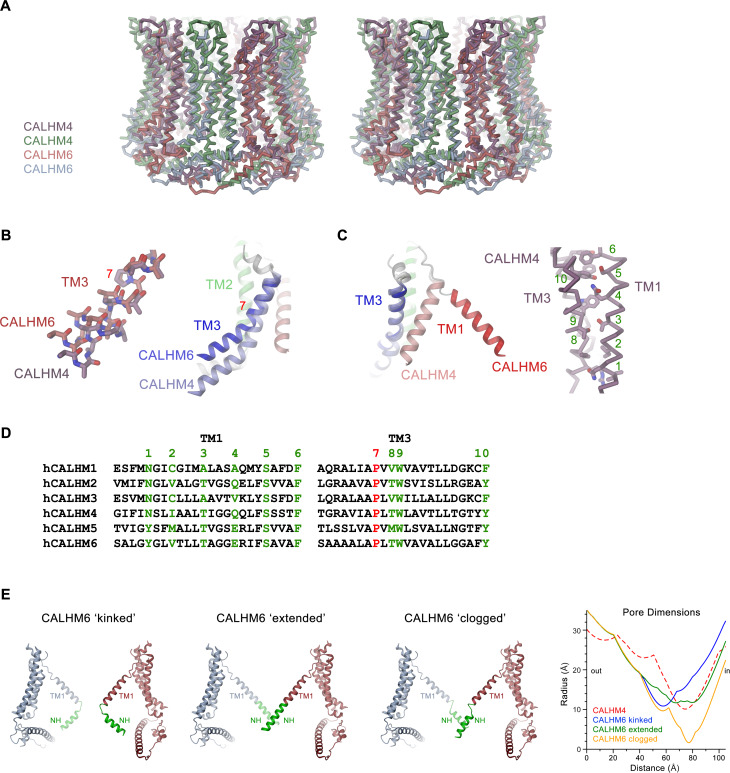
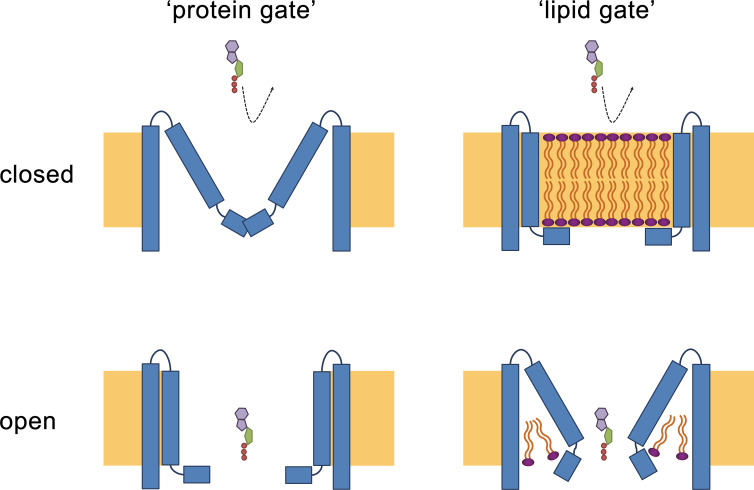
The transport of substances across the placenta is essential for the development of the fetus. Here, we were interested in the role of channels of the calcium homeostasis modulator (CALHM) family in the human placenta. By transcript analysis, we found the paralogs CALHM2, 4, and 6 to be highly expressed in this organ and upregulated during trophoblast differentiation. Based on electrophysiology, we observed that activation of these paralogs differs from the voltage- and calcium-gated channel CALHM1. Cryo-EM structures of CALHM4 display decameric and undecameric assemblies with large cylindrical pore, while in CALHM6 a conformational change has converted the pore shape into a conus that narrows at the intracellular side, thus describing distinct functional states of the channel. The pore geometry alters the distribution of lipids, which occupy the cylindrical pore of CALHM4 in a bilayer-like arrangement whereas they have redistributed in the conical pore of CALHM6 with potential functional consequences.
物质跨胎盘转运对胎儿发育至关重要。在这里,我们研究了钙稳态调节剂(CALHM)家族通道在人胎盘中的作用。通过转录分析,我们发现同源物 CALHM2、4 和 6 在该器官中高度表达,并在滋养细胞分化过程中上调。基于电生理学,我们观察到这些同源物的激活不同于电压和钙门控通道 CALHM1。CALHM4 的冷冻电镜结构显示出具有大圆柱形孔的十聚体和十一聚体组装,而在 CALHM6 中,构象变化将孔形状转换为圆锥体,在细胞内侧面变窄,从而描述了通道的不同功能状态。孔几何形状改变了脂质的分布,CALHM4 的圆柱形孔中脂质呈双层排列,而在 CALHM6 的圆锥形孔中脂质重新分布,可能具有潜在的功能后果。