Gebregiwergis G T, Sørensen Anders C, Henryon Mark, Meuwissen Theo
Department of Animal and Aquaculture Sciences, Norwegian University of Life Sciences, Ås, Norway.
Department of Molecular Biology and Genetics, Aarhus University, Aarhus, Denmark.
Front Genet. 2020 Apr 22;11:345. doi: 10.3389/fgene.2020.00345. eCollection 2020.
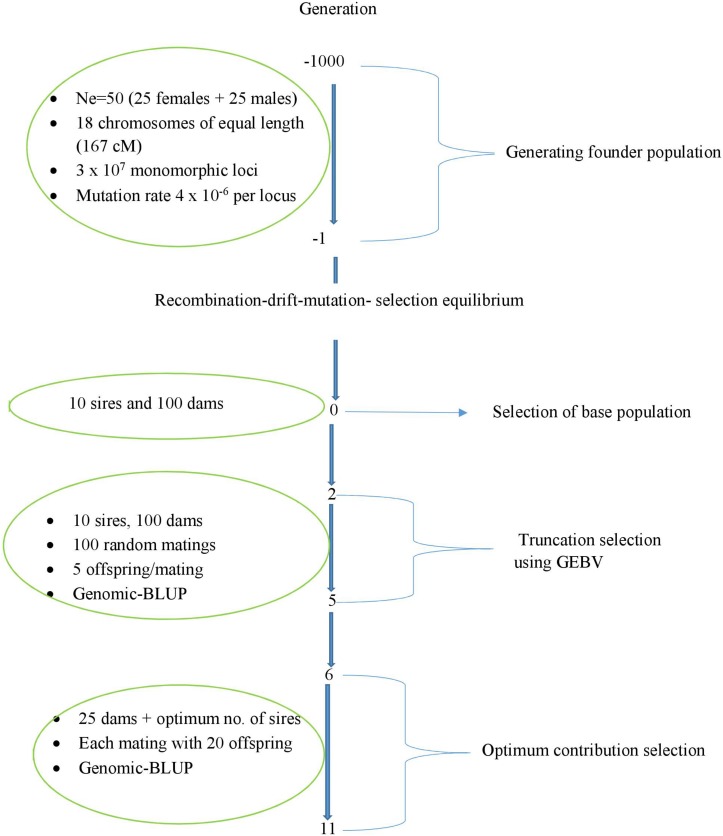
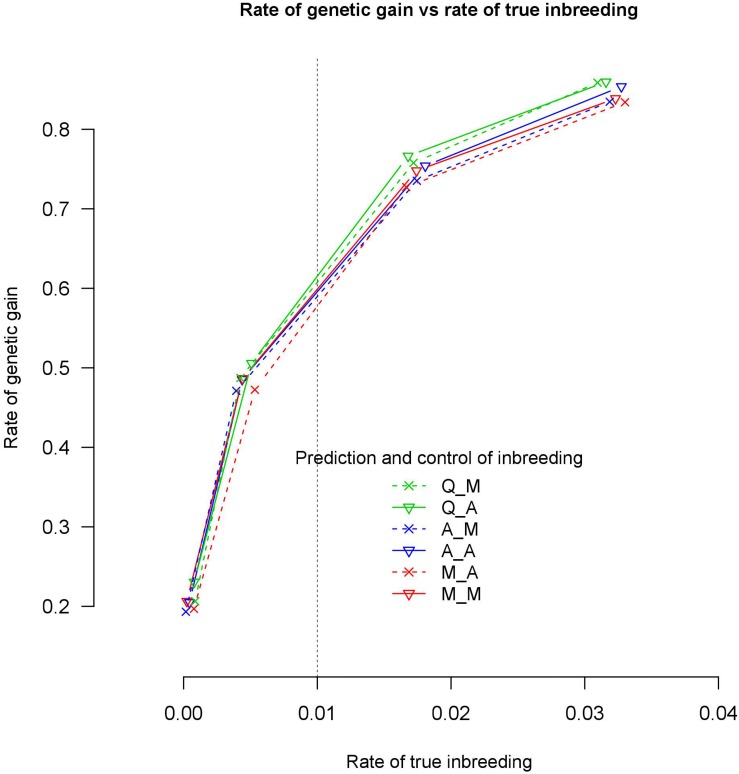
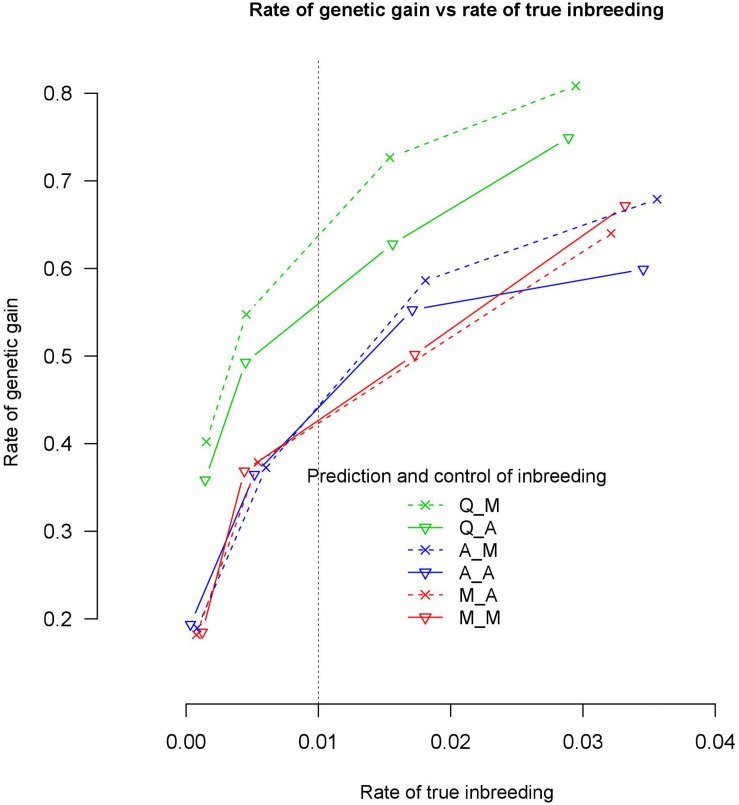
We tested the consequences of using alternative genomic relationship matrices to predict genomic breeding values (GEBVs) and control of coancestry in optimum contribution selection, where the relationship matrix used to calculate GEBVs was not necessarily the same as that used to control coancestry. A stochastic simulation study was carried out to investigate genetic gain and true genomic inbreeding in breeding schemes that applied genomic optimum contribution selection (GOCS) with different genomic relationship matrices. Three genomic-relationship matrices were used to predict the GEBVs based on three information sources: markers ( ), QTL ( ), and markers and QTL ( ). Strictly, is not possible to implement in practice since we do not know the quantitative trait loci (QTL) positions, but more and more information is becoming available especially about the largest QTL. Two genomic-relationship matrices were used to control coancestry: and . Three genetic architectures were simulated: with 7702, 1000, and 500 QTLs together with 54,218 markers. Selection was for a single trait with heritability 0.2. All selection candidates were phenotyped and genotyped before selection. With 7702 QTL, there were no significant differences in rates of genetic gain at the same rate of true inbreeding using different genomic relationship matrices in GOCS. However, as the number of QTLs was reduced to 1000, prediction of GEBVs using a genomic relationship matrix constructed based on and control of coancestry using realized 29.7% higher genetic gain than using for both prediction and control of coancestry. Forty-three percent of this increased rate of genetic gain was due to increased accuracies of GEBVs. These findings indicate that with large numbers of QTL, it is not critical what information, i.e., markers or QTL, is used to construct genomic-relationship matrices. However, it becomes critical with small numbers of QTL. This highlights the importance of using genomic-relationship matrices that focus on QTL regions for GEBV estimation when the number of QTL is small in GOCS. Relationships used to control coancestry are preferably based on marker data.
我们测试了在最优贡献选择中使用替代基因组关系矩阵来预测基因组育种值(GEBVs)和控制同祖系数的后果,其中用于计算GEBVs的关系矩阵不一定与用于控制同祖系数的关系矩阵相同。开展了一项随机模拟研究,以调查在应用不同基因组关系矩阵的基因组最优贡献选择(GOCS)育种方案中的遗传进展和真实基因组近亲繁殖情况。基于三种信息来源使用了三种基因组关系矩阵来预测GEBVs:标记( )、QTL( )以及标记和QTL( )。严格来说, 在实际中无法实施,因为我们不知道数量性状位点(QTL)的位置,但越来越多的信息尤其是关于最大QTL的信息变得可用。使用了两种基因组关系矩阵来控制同祖系数: 和 。模拟了三种遗传结构:分别有7702个、1000个和500个QTL以及54,218个标记。针对遗传力为0.2的单一性状进行选择。所有选择候选个体在选择前都进行了表型测定和基因分型。对于有7702个QTL的情况,在GOCS中使用不同基因组关系矩阵且真实近亲繁殖率相同时,遗传进展速率没有显著差异。然而,当QTL数量减少到1000个时,使用基于 构建的基因组关系矩阵预测GEBVs并使用 控制同祖系数,相比于使用 进行GEBVs预测和同祖系数控制,实现了高29.7%的遗传进展。这一遗传进展增加率的43%归因于GEBVs准确性的提高。这些发现表明,当有大量QTL时,使用何种信息(即标记或QTL)来构建基因组关系矩阵并不关键。然而,当QTL数量较少时,这就变得至关重要。这凸显了在GOCS中QTL数量较少时,使用专注于QTL区域的基因组关系矩阵进行GEBV估计的重要性。用于控制同祖系数的关系最好基于标记数据。