Department of Chemistry, East Carolina University, Greenville, North Carolina 27858, United States.
Department of Chemistry and Francis Bitter Magnet Laboratory, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, United States.
Biochemistry. 2020 Jun 30;59(25):2319-2327. doi: 10.1021/acs.biochem.0c00079. Epub 2020 Jun 14.
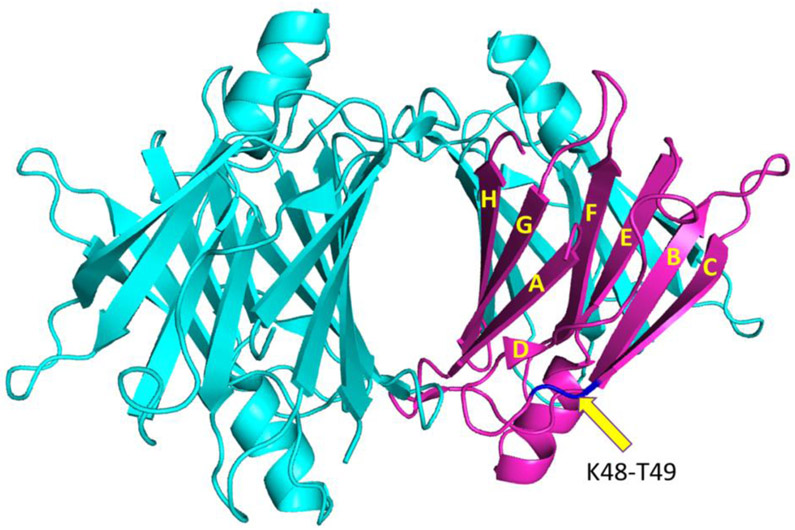
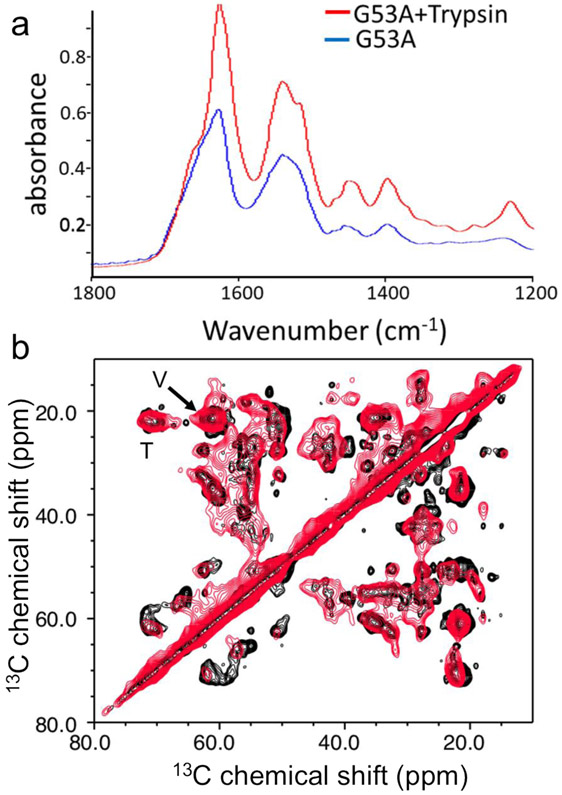
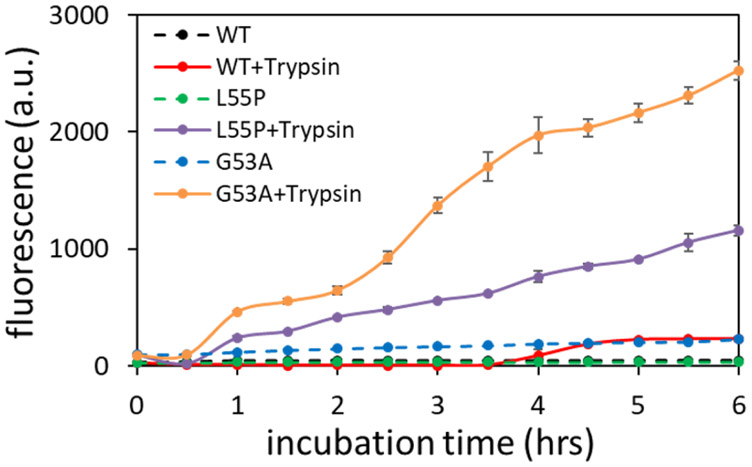
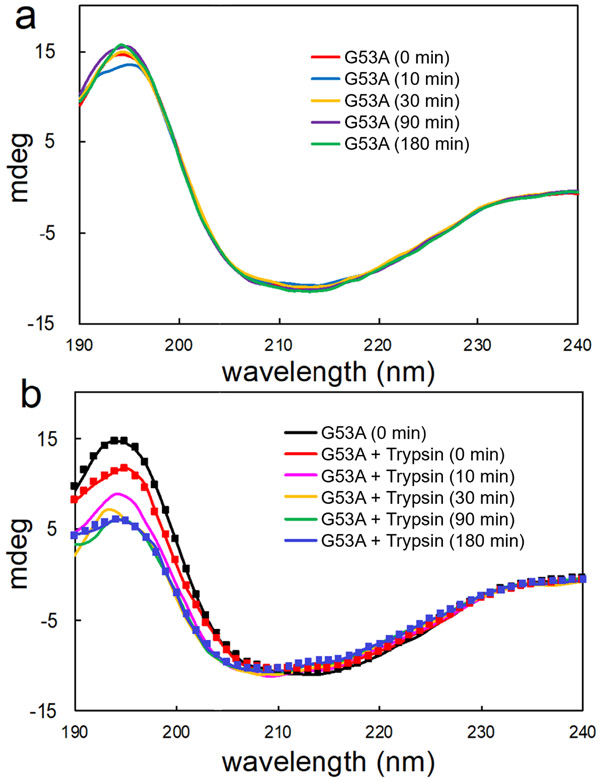
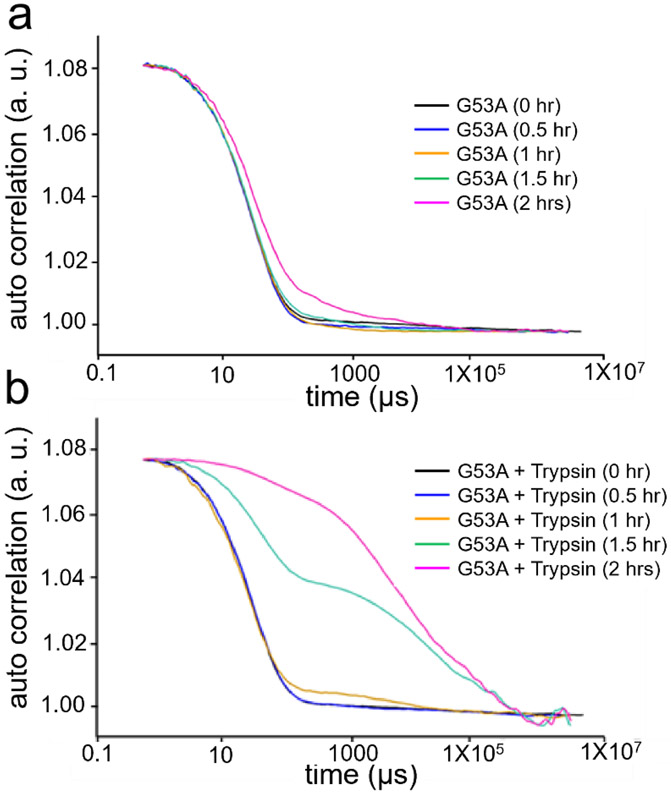
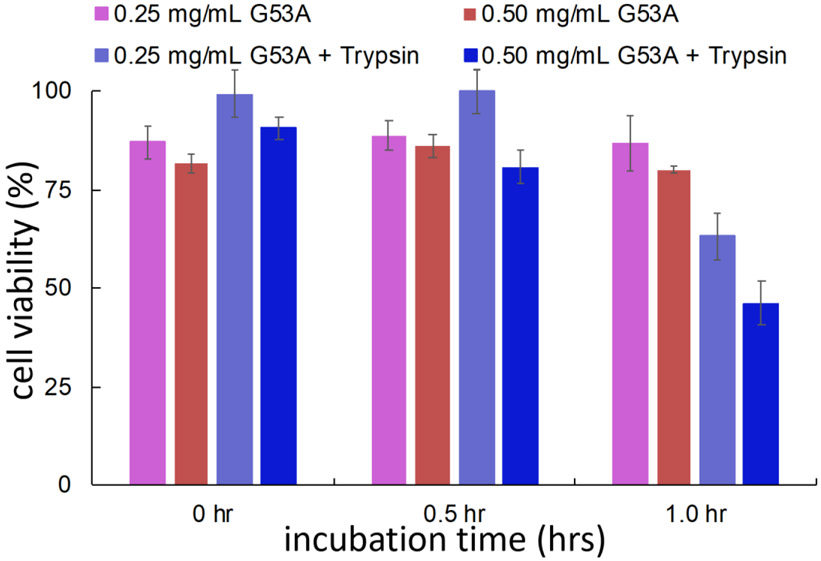
Amyloid formation of full-length TTR involves dissociation of the native tetramers into misfolded monomers that self-assemble into amyloid. In addition to the full-length TTR, C-terminal fragments including residues 49-127 were also observed , implying the presence of additional misfolding pathways. It was previously proposed that a proteolytic cleavage might lead to the formation of the C-terminal fragment TTR amyloid. Here, we report mechanistic studies of misfolding and aggregation of a TTR variant (G53A) in the absence and presence of a serine protease. A proteolytic cleavage of G53A in the CD loop (K48 and T49) with agitation promoted TTR misfolding and aggregation, suggesting that the proteolytic cleavage may lead to the aggregation of the C-terminal fragment (residues 49-127). To gain more detailed insights into TTR misfolding promoted by proteolytic cleavage, we investigated structural changes in G53A TTR in the presence and absence of trypsin. Our combined biophysical analyses revealed that the proteolytic cleavage accelerated the formation of spherical small oligomers, which exhibited cytotoxic activities. However, the truncated TTR appeared to maintain native-like structures, rather than the C-terminal fragment (residues 49-127) being released and unfolded from the native state. In addition, our solid-state nuclear magnetic resonance and Fourier transform infrared structural studies showed that the two aggregates derived from the full-length and cleaved TTR exhibited nearly identical molecular structural features, suggesting that the proteolytic cleavage in the CD loop destabilizes the native tetrameric structure and accelerates oligomer formation through a common TTR misfolding and aggregation mechanism rather than through a distinct molecular mechanism.
全长 TTR 的淀粉样蛋白形成涉及到天然四聚体的解离,形成错误折叠的单体,这些单体自组装成淀粉样蛋白。除了全长 TTR 外,还观察到包括残基 49-127 的 C 端片段,这表明存在额外的错误折叠途径。先前有人提出,蛋白水解切割可能导致 C 端片段 TTR 淀粉样蛋白的形成。在这里,我们报告了在缺乏和存在丝氨酸蛋白酶的情况下,TTR 变体(G53A)错误折叠和聚集的机制研究。在搅拌下,G53A 在 CD 环(K48 和 T49)上的蛋白水解切割促进了 TTR 的错误折叠和聚集,表明蛋白水解切割可能导致 C 端片段(残基 49-127)的聚集。为了更详细地了解蛋白水解切割促进的 TTR 错误折叠,我们研究了在有和没有胰蛋白酶存在的情况下 G53A TTR 的结构变化。我们的综合生物物理分析表明,蛋白水解切割加速了球形小寡聚物的形成,这些寡聚物表现出细胞毒性活性。然而,截断的 TTR 似乎保持了类似天然的结构,而不是 C 端片段(残基 49-127)从天然状态中释放和展开。此外,我们的固态核磁共振和傅里叶变换红外结构研究表明,来自全长和切割 TTR 的两种聚集体表现出几乎相同的分子结构特征,这表明 CD 环中的蛋白水解切割破坏了天然四聚体结构,并通过共同的 TTR 错误折叠和聚集机制加速寡聚体形成,而不是通过独特的分子机制。