Department of Chemistry, University of Massachusetts, Amherst, United States.
Department of Biomedical Engineering, Center for the Investigation of Membrane Excitability Disorders, Cardiac Bioelectricity and Arrhythmia Center, Washington University, St Louis, United States.
Elife. 2020 Jun 29;9:e55571. doi: 10.7554/eLife.55571.
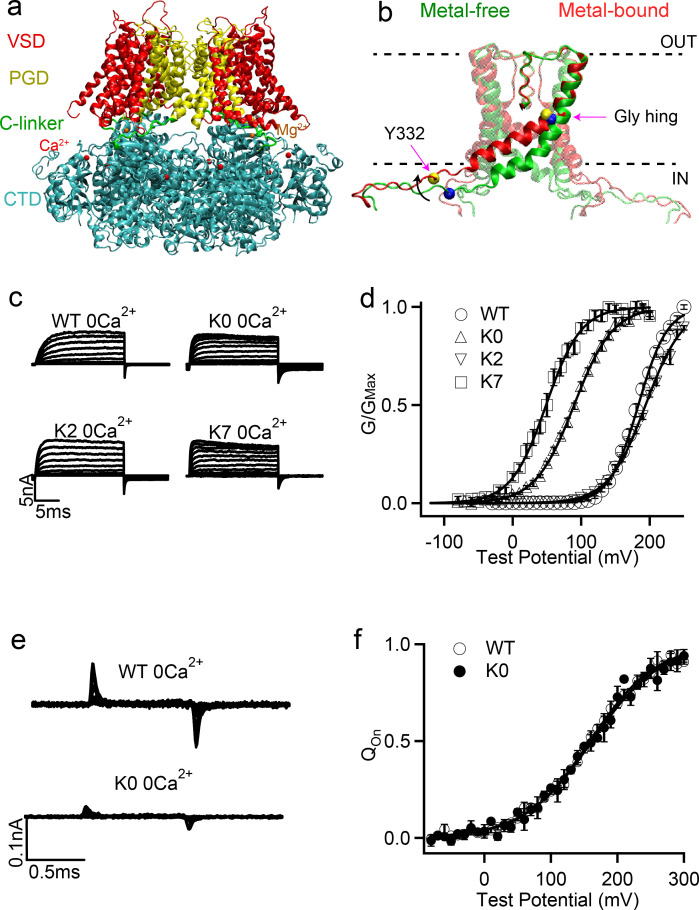
Large-conductance potassium (BK) channels are transmembrane (TM) proteins that can be synergistically and independently activated by membrane voltage and intracellular Ca. The only covalent connection between the cytosolic Ca sensing domain and the TM pore and voltage sensing domains is a 15-residue 'C-linker'. To determine the linker's role in human BK activation, we designed a series of linker sequence scrambling mutants to suppress potential complex interplay of specific interactions with the rest of the protein. The results revealed a surprising sensitivity of BK activation to the linker sequence. Combining atomistic simulations and further mutagenesis experiments, we demonstrated that nonspecific interactions of the linker with membrane alone could directly modulate BK activation. The C-linker thus plays more direct roles in mediating allosteric coupling between BK domains than previously assumed. Our results suggest that covalent linkers could directly modulate TM protein function and should be considered an integral component of the sensing apparatus.
大电导钙激活钾(BK)通道是跨膜(TM)蛋白,可被膜电压和细胞内 Ca 协同且独立地激活。细胞溶质 Ca 感应域和 TM 孔及电压感应域之间唯一的共价连接是一个 15 个残基的“C 连接子”。为了确定连接子在人 BK 激活中的作用,我们设计了一系列连接子序列改组突变体,以抑制与蛋白质其余部分的特定相互作用的潜在复杂相互作用。结果显示,BK 激活对连接子序列具有惊人的敏感性。通过原子模拟和进一步的突变实验,我们证明了连接子与膜的非特异性相互作用本身就可以直接调节 BK 的激活。因此,与之前的假设相比,C 连接子在介导 BK 结构域的变构偶联中发挥着更直接的作用。我们的研究结果表明,共价连接子可以直接调节 TM 蛋白的功能,应被视为感应装置的一个组成部分。