Tel-Zur Noemi, Mouyal Joseph, Zurgil Udi, Mizrahi Yosef
The French Associates Institute for Agriculture and Biotechnology of Drylands, The Jacob Blaustein Institutes for Desert Research, Ben-Gurion University of the Negev, Beer-Sheva, Israel.
Department of Life Sciences, Ben-Gurion University of the Negev, Beer-Sheva, Israel.
Front Plant Sci. 2020 Jun 26;11:954. doi: 10.3389/fpls.2020.00954. eCollection 2020.
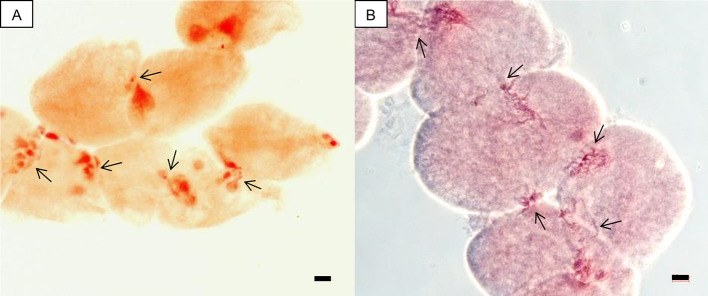
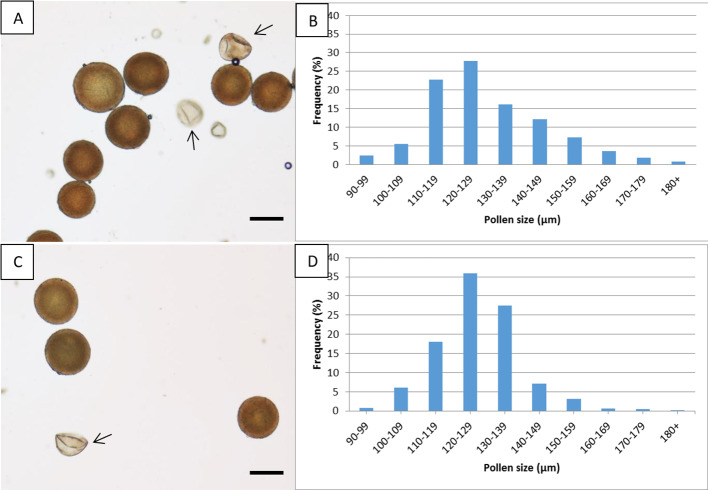
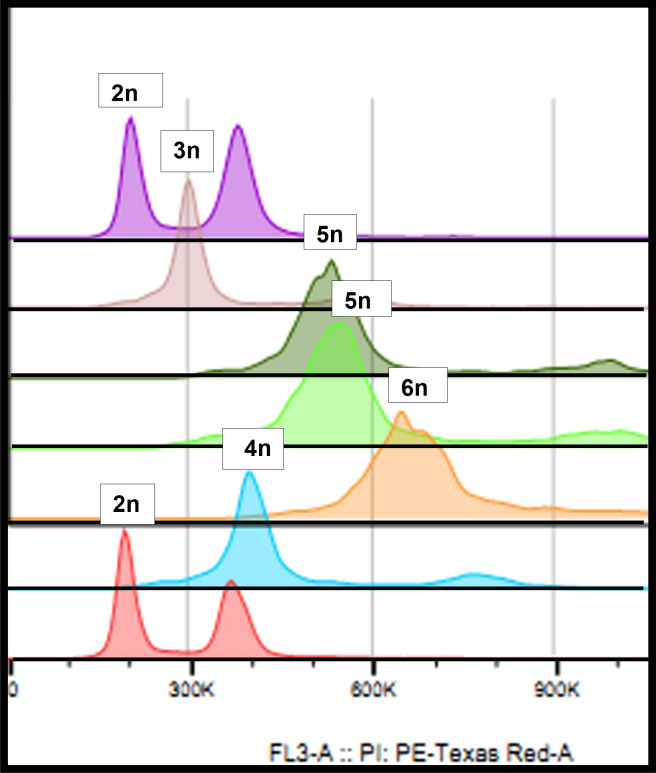
Polyploidy-or chromosome doubling-plays a significant role in plant speciation and evolution. Much of the existing evidence indicates that fusion of unreduced (or 2) gametes is the major pathway responsible for polyploid formation. In the early 1900s, a theory was put forward that the mechanism of "hybridization followed by chromosome doubling" would enable the survival and development of the hybrid zygote by providing each chromosome with a homolog with which to pair. However, to date there is only scant empirical evidence supporting this theory. In our previous study, interspecific-interploid crosses between the tetraploid as the female parent, and the diploid as the male parent, yielded only allopentaploids, allohexaploids, and 5-and 6-aneuploids instead of the expected allotriploids. No viable hybrids were obtained from the reciprocal cross. Since underwent normal meiosis with regular pairing in the pollen mother cells and only reduced pollen grains were observed, the allohexaploids obtained supported the concept of "chromosome doubling." In this work, we report ploidy level, fruit morphology, and pollen viability and diameter in a group of putative hybrids obtained from an embryo rescue procedure following controlled × crosses, with the aim to elucidate, for the first time, the timing and developmental stage of the chromosome doubling. As in our previous report, no triploids were obtained, but tetraploids, pentaploids, hexaploids, and 5- and 6-aneuploids were found in the regenerated plants. The tetraploids exhibited the morphological features of the maternal parent and could not be considered true hybrids. Based on our previous studies, we can assume that the pentaploids were a result of a fertilization event between one unreduced (2) female gamete from the tetraploid and a normal () haploid male gamete from . All the allohexaploids obtained from the embryo rescue technique where those that regenerated from fertilized ovules 10 days after pollination (at the pro-embryo stage), showing that the chromosome doubling event occurred at a very early development stage, i.e., at the zygote stage or shortly after zygote formation. These allohexaploids thus constitute empirical evidence of "hybridization followed by chromosome doubling."
多倍体现象,即染色体加倍,在植物物种形成和进化过程中发挥着重要作用。现有大量证据表明,未减数(或2n)配子的融合是多倍体形成的主要途径。20世纪初,有人提出“杂交后染色体加倍”的机制能够通过为每条染色体提供一个同源配对染色体,使杂交合子得以存活和发育。然而,迄今为止,仅有少量实证证据支持这一理论。在我们之前的研究中,以四倍体为母本、二倍体为父本进行种间倍间杂交,结果只产生了异源五倍体、异源六倍体以及5 - 体和6 - 体非整倍体,而非预期的异源三倍体。正反交均未获得可育杂种。由于在花粉母细胞中正常减数分裂且配对正常,仅观察到减数花粉粒,所以获得的异源六倍体支持了“染色体加倍”的概念。在这项研究中,我们报告了一组通过控制×杂交后胚胎拯救程序获得的假定杂种的倍性水平、果实形态、花粉活力和直径,旨在首次阐明染色体加倍的时间和发育阶段。与我们之前的报告一样,未获得三倍体,但在再生植株中发现了四倍体、五倍体、六倍体以及5 - 体和6 - 体非整倍体。四倍体表现出母本的形态特征,不能视为真正的杂种。基于我们之前的研究,我们可以假定五倍体是四倍体的一个未减数(2n)雌配子与的一个正常(n)单倍体雄配子受精的结果。通过胚胎拯救技术获得的所有异源六倍体均来自授粉10天后(原胚阶段)受精胚珠再生的植株,这表明染色体加倍事件发生在非常早期的发育阶段,即合子阶段或合子形成后不久。因此,这些异源六倍体构成了“杂交后染色体加倍”的实证证据。