Department of Molecular, Cell and Developmental Biology, University of California, Santa Cruz, Santa Cruz, CA 95064.
Mol Biol Cell. 2020 Sep 15;31(20):2219-2233. doi: 10.1091/mbc.E20-05-0310. Epub 2020 Jul 22.
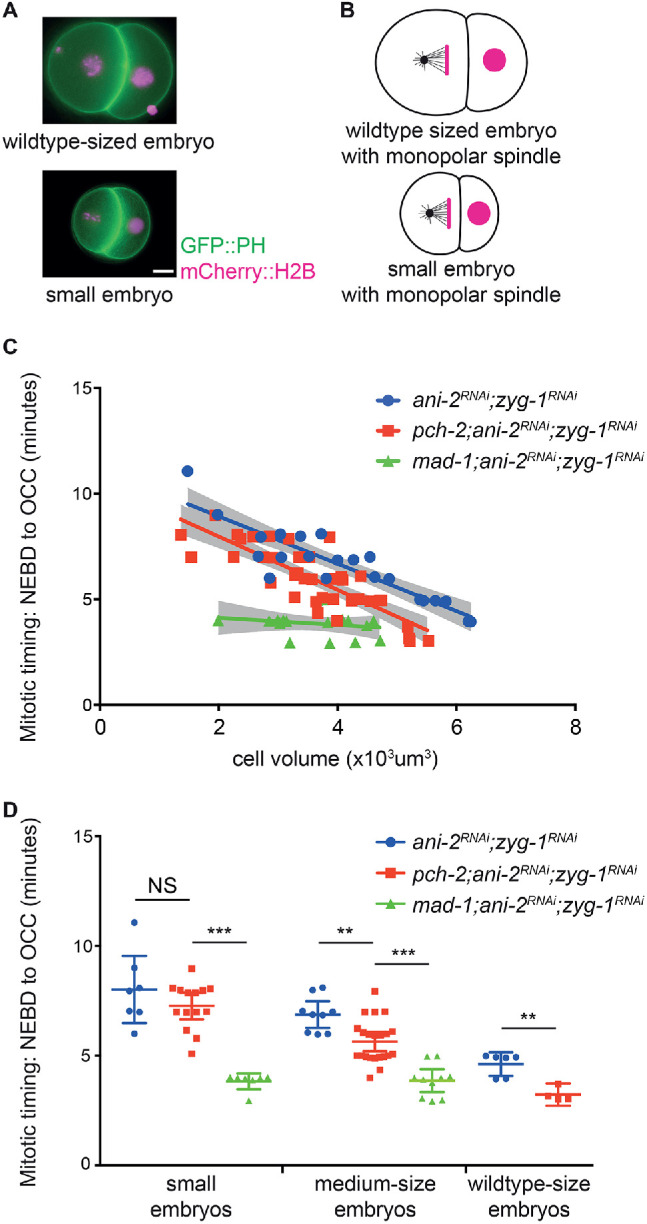
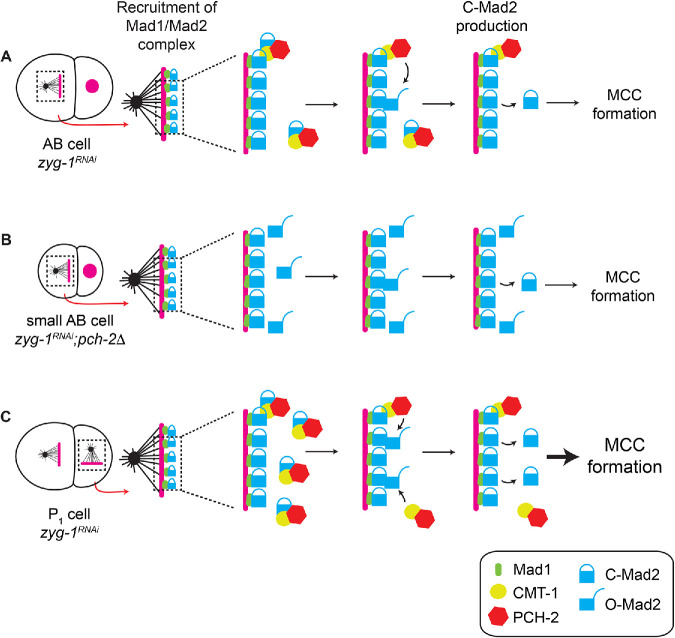
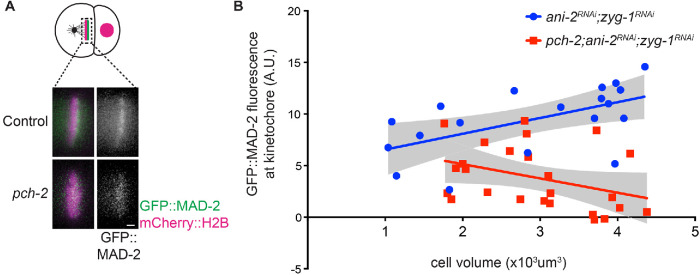
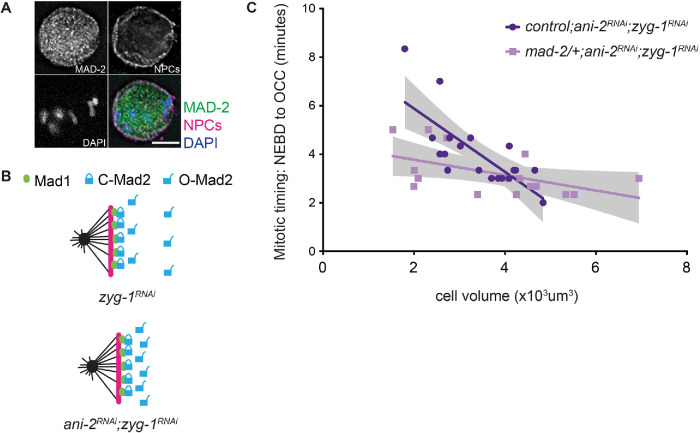
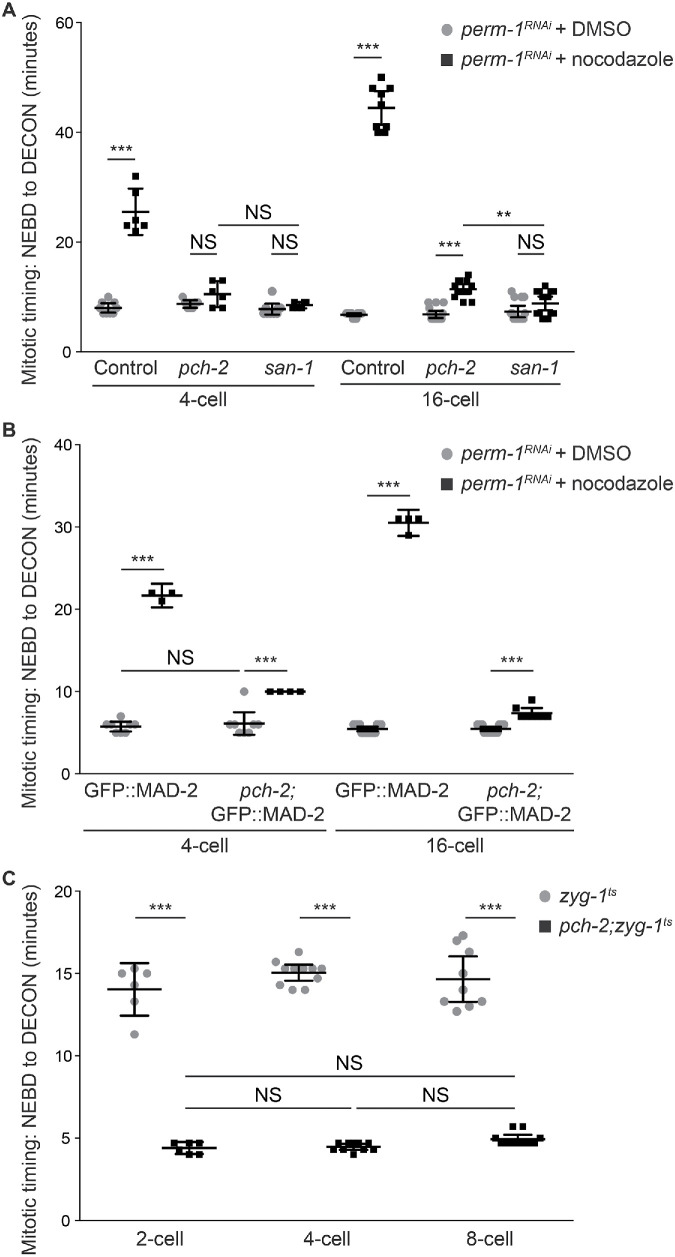
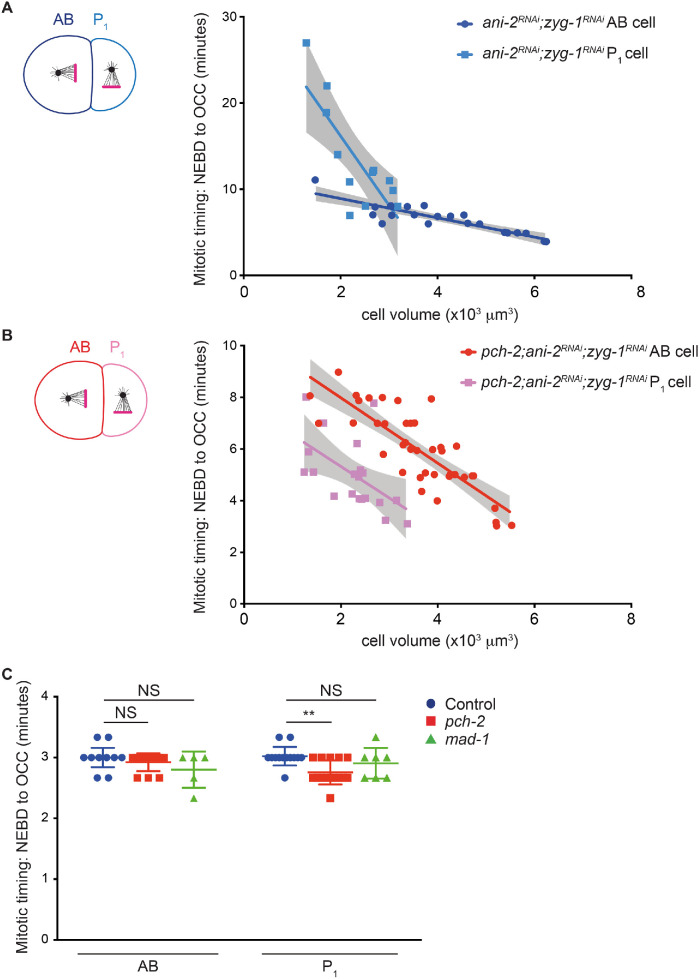
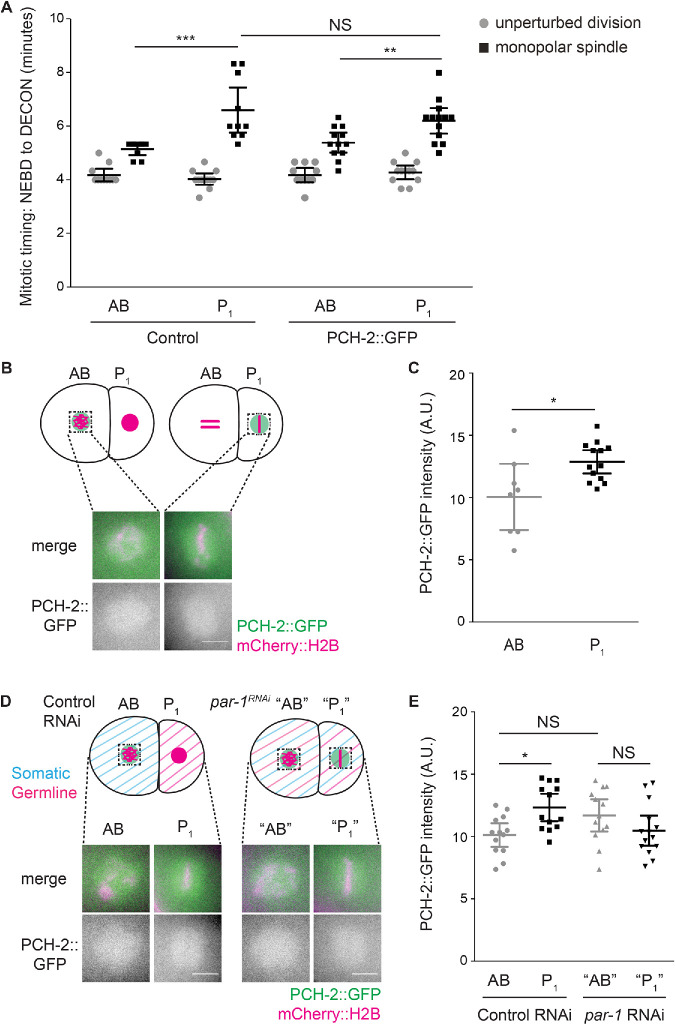
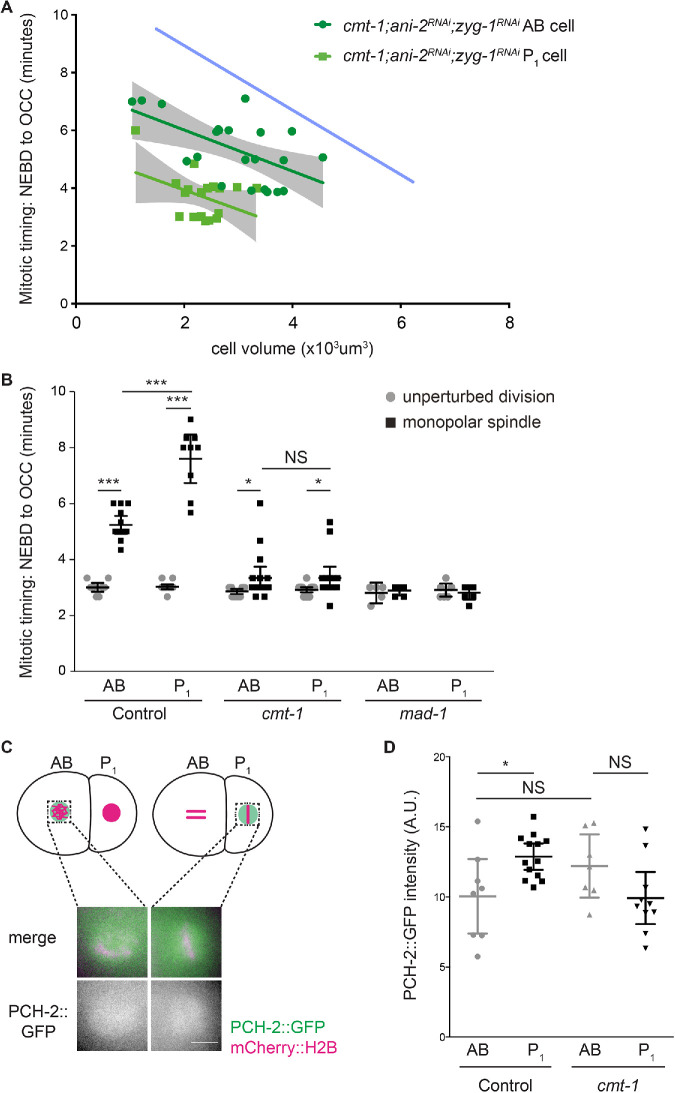
Spindle checkpoint strength is dictated by the number of unattached kinetochores, cell volume, and cell fate. We show that the conserved AAA-ATPase PCH-2/TRIP13, which remodels the checkpoint effector Mad2 from an active conformation to an inactive one, controls checkpoint strength in . Having previously established that this function is required for spindle checkpoint activation, we demonstrate that in cells genetically manipulated to decrease in cell volume, PCH-2 is no longer required for the spindle checkpoint or recruitment of Mad2 at unattached kinetochores. This role is not limited to large cells: the stronger checkpoint in germline precursor cells also depends on PCH-2. PCH-2 is enriched in germline precursor cells, and this enrichment relies on conserved factors that induce asymmetry in the early embryo. Finally, the stronger checkpoint in germline precursor cells is regulated by CMT-1, the ortholog of p31, which is required for both PCH-2's localization to unattached kinetochores and its enrichment in germline precursor cells. Thus, PCH-2, likely by regulating the availability of inactive Mad2 at and near unattached kinetochores, governs checkpoint strength. This requirement may be particularly relevant in oocytes and early embryos enlarged for developmental competence, cells that divide in syncytial tissues, and immortal germline cells
纺锤体检验点的强度取决于游离的动粒数量、细胞体积和细胞命运。我们发现,保守的 AAA-ATP 酶 PCH-2/TRIP13 可以将检验点效应物 Mad2 从活性构象转变为非活性构象,从而控制. 的检验点强度。我们之前已经证明,该功能对于纺锤体检验点的激活是必需的,我们证明在通过遗传操作降低细胞体积的细胞中,PCH-2 不再需要用于纺锤体检验点或在未连接的动粒处募集 Mad2。这个作用不仅限于大细胞:生殖细胞前体细胞中更强的检验点也依赖于 PCH-2。PCH-2 在生殖细胞前体细胞中富集,这种富集依赖于诱导早期胚胎不对称的保守因素。最后,生殖细胞前体细胞中更强的检验点受 CMT-1 的调节,CMT-1 是 p31 的同源物,它既需要 PCH-2 定位于未连接的动粒,也需要其在生殖细胞前体细胞中的富集。因此,PCH-2 可能通过调节游离动粒处和附近未连接的动粒上失活 Mad2 的可用性来控制检验点强度。这个要求在为发育能力而增大的卵母细胞和早期胚胎、在合胞组织中分裂的细胞以及永生的生殖细胞中可能特别相关。