Institute for Cell Biology, Medical University of Innsbruck, Innsbruck, Austria.
ADSI - Austrian Drug Screening Institute GmbH, Innsbruck, Austria.
Elife. 2020 Aug 3;9:e58246. doi: 10.7554/eLife.58246.
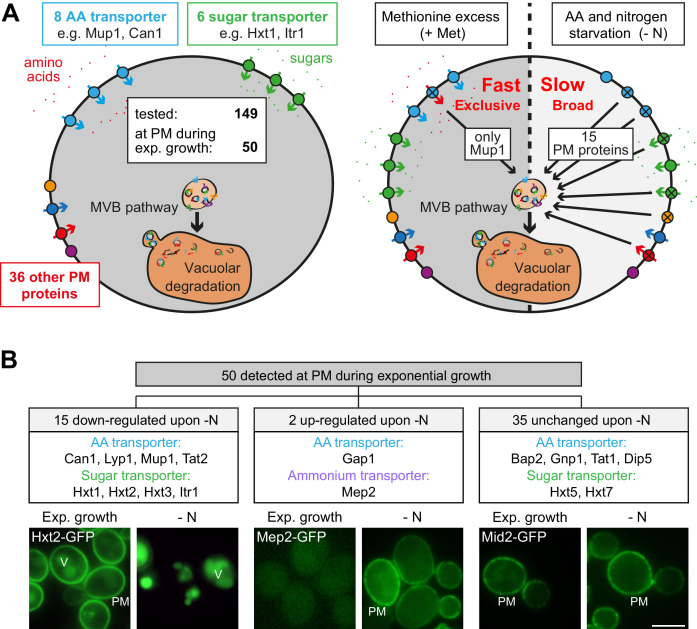
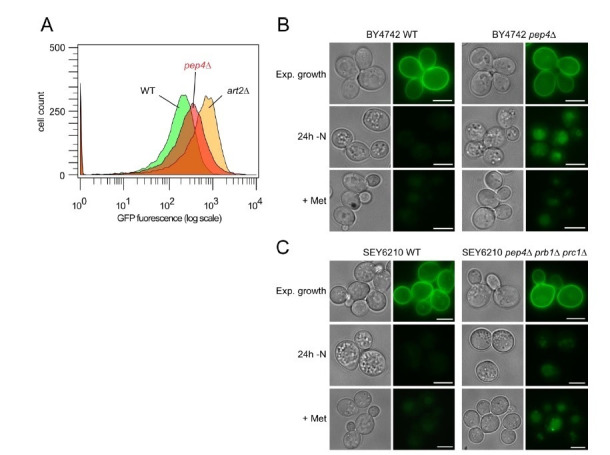
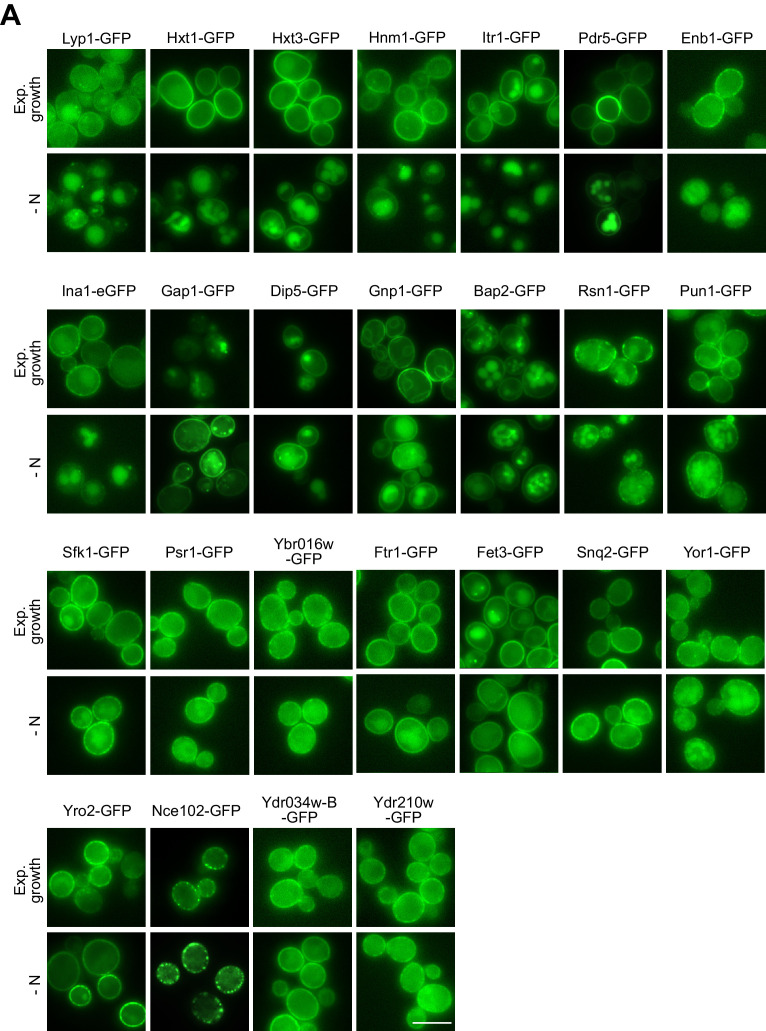
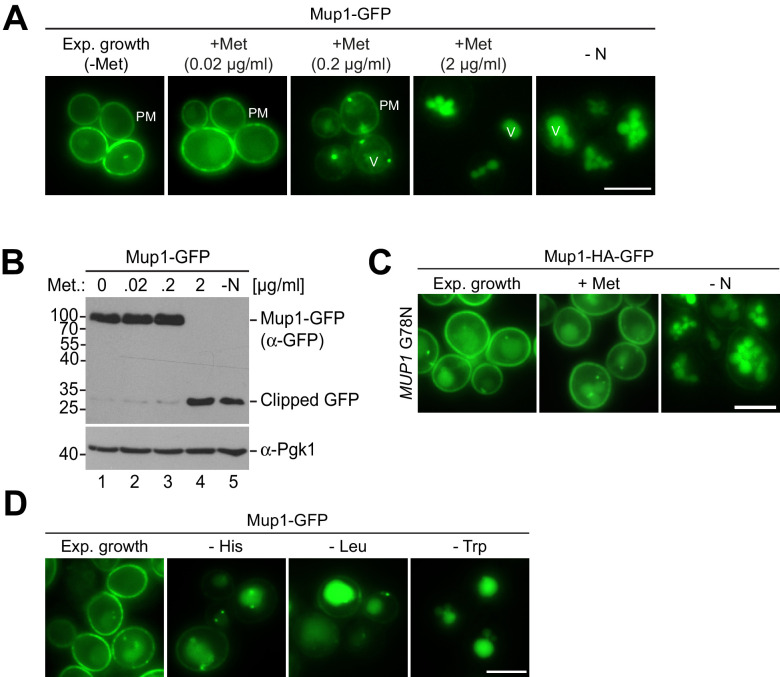
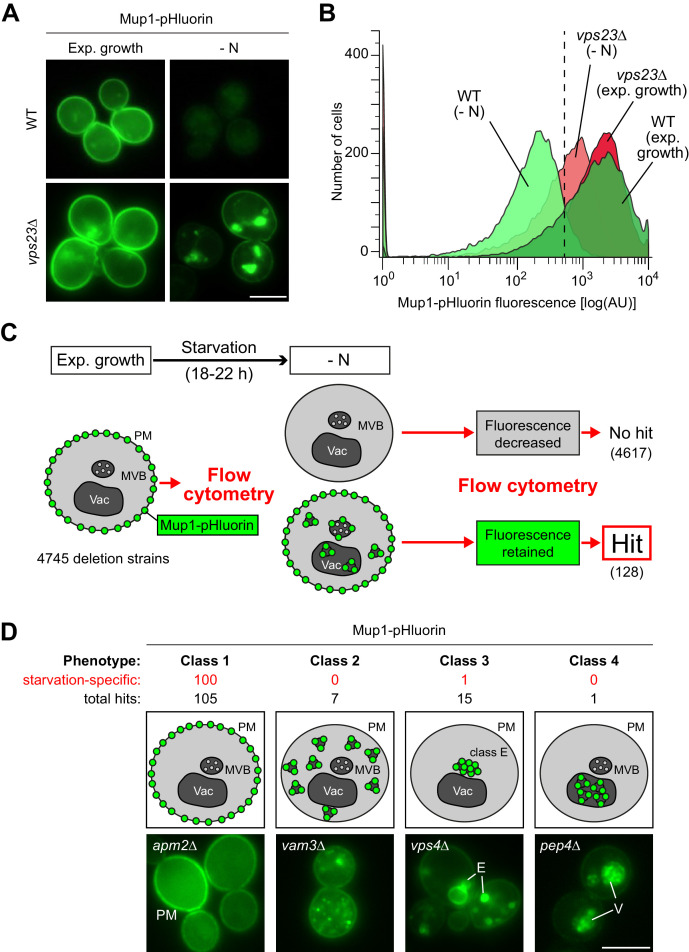
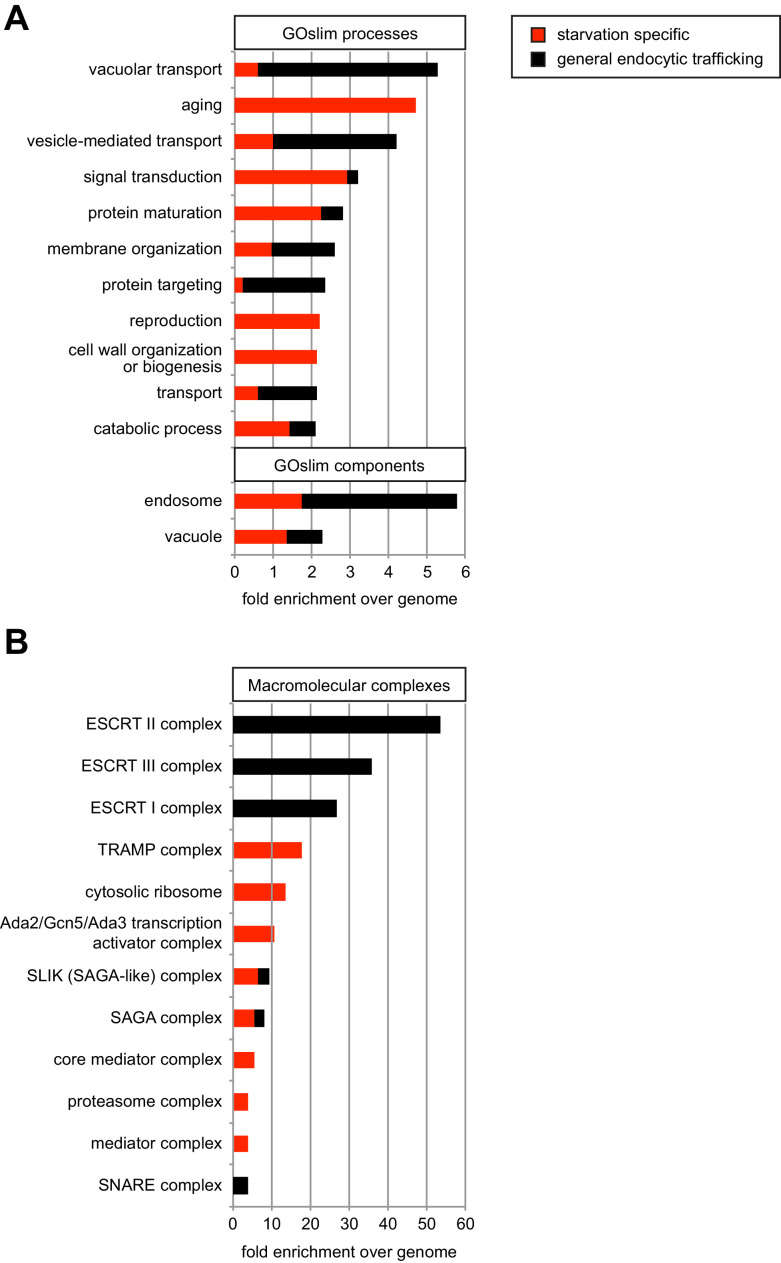
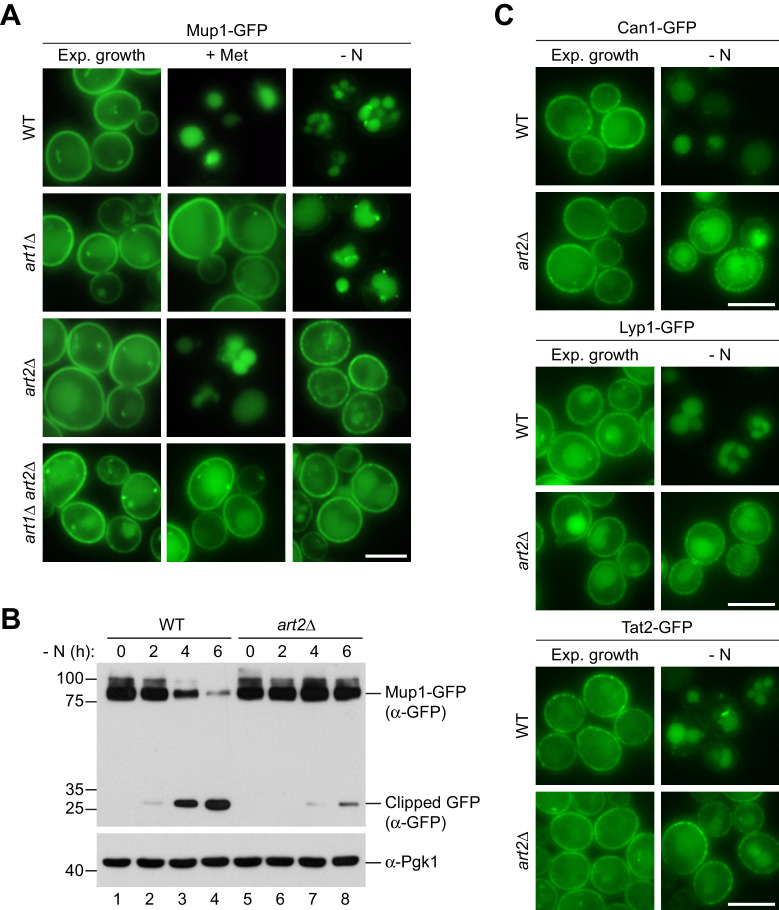
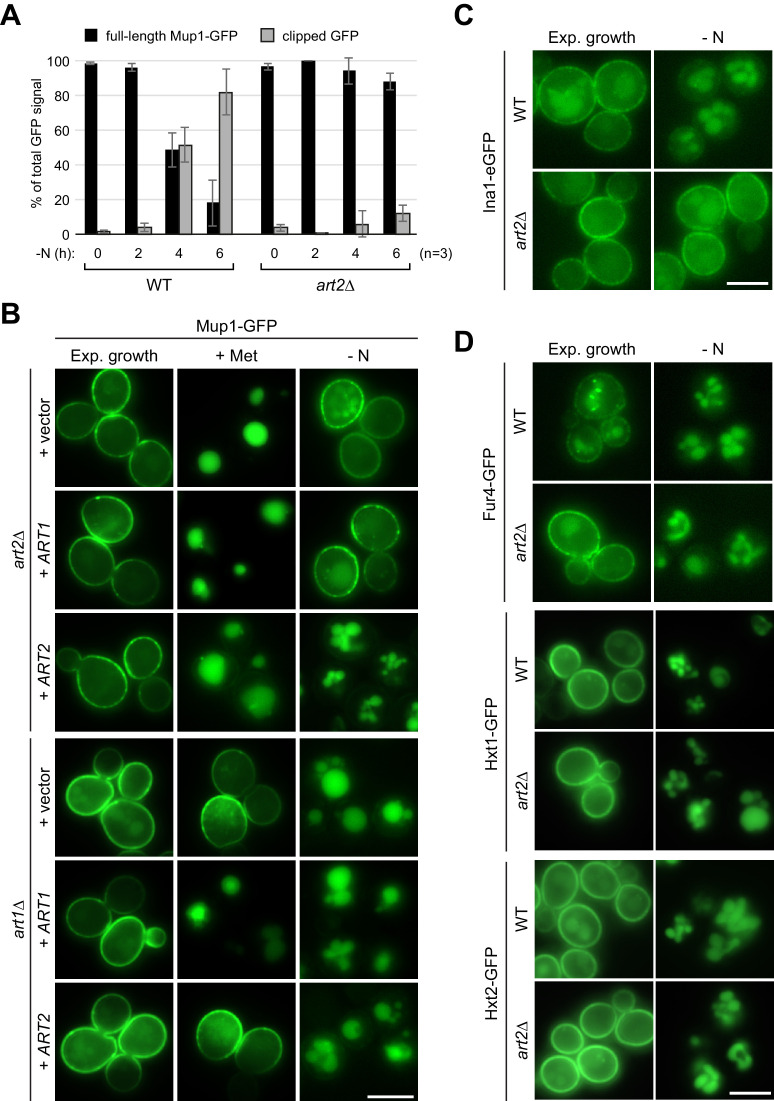
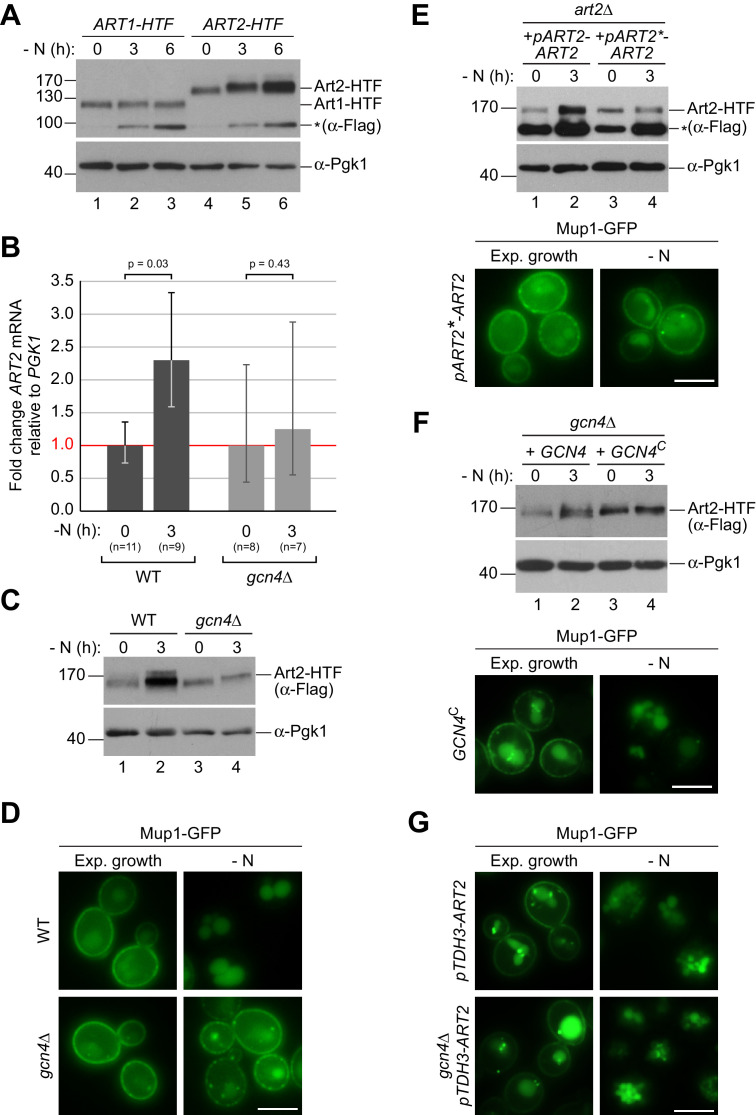
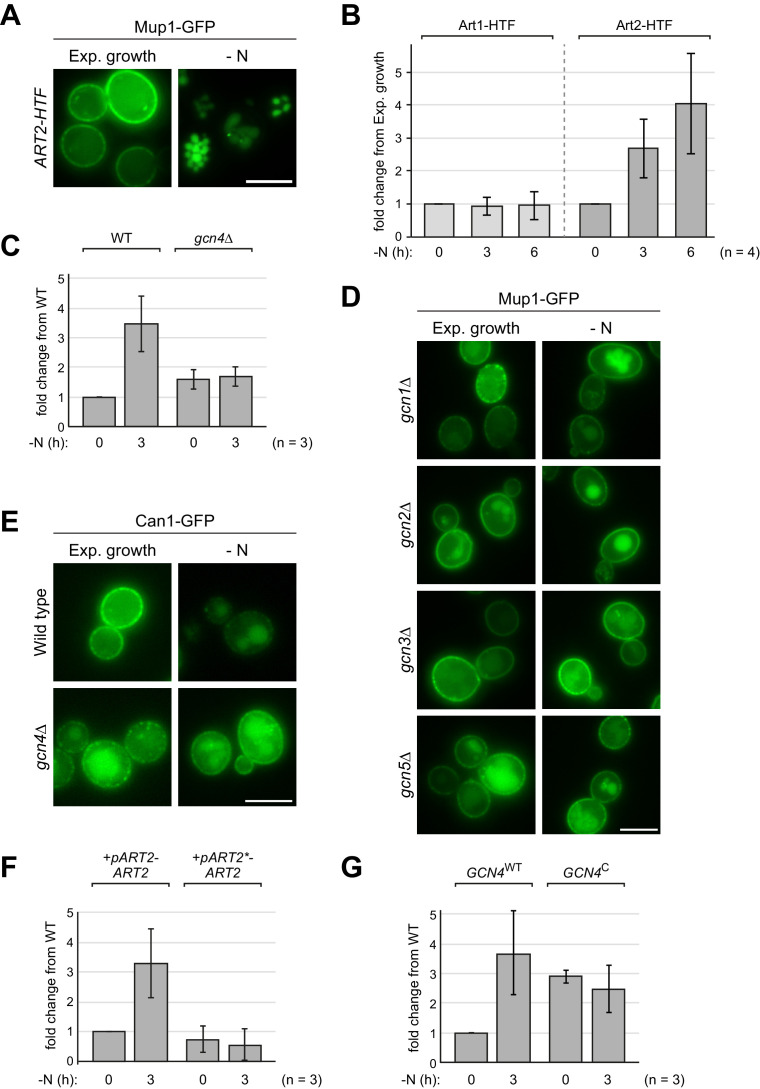
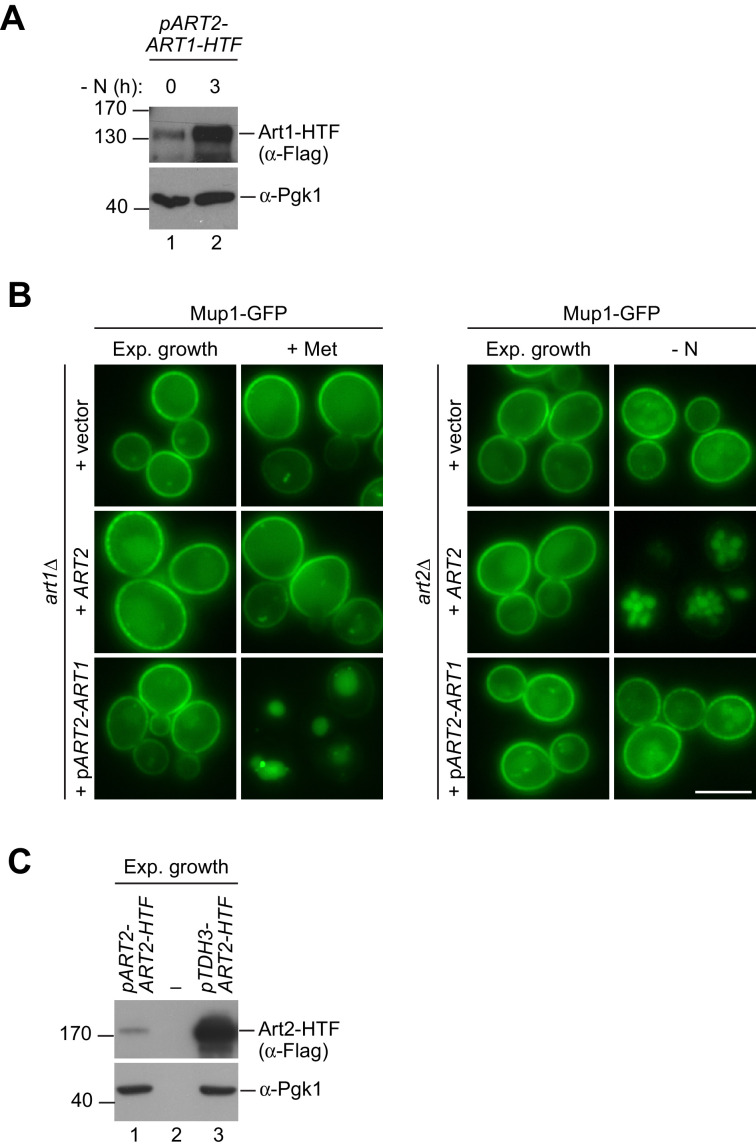
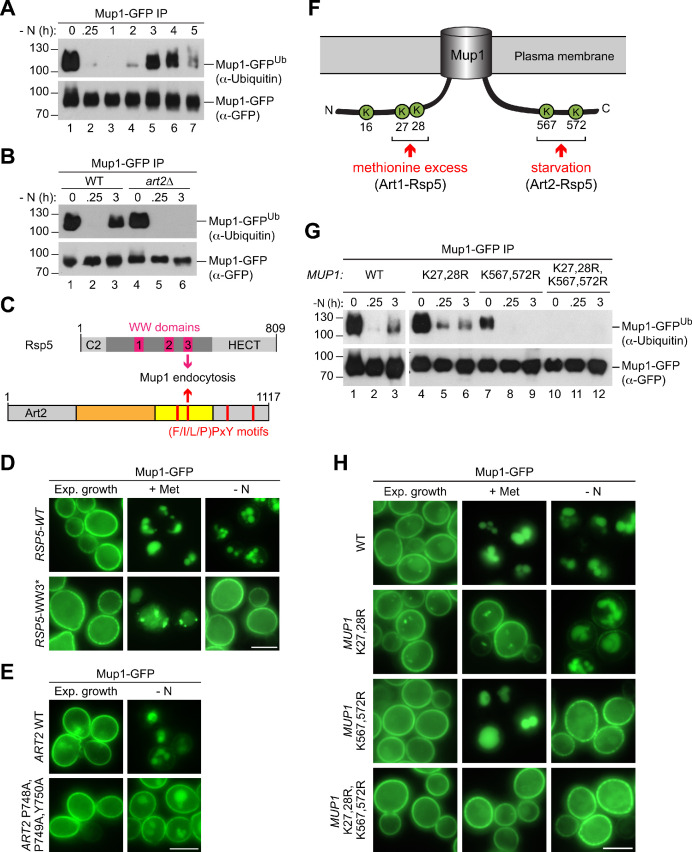
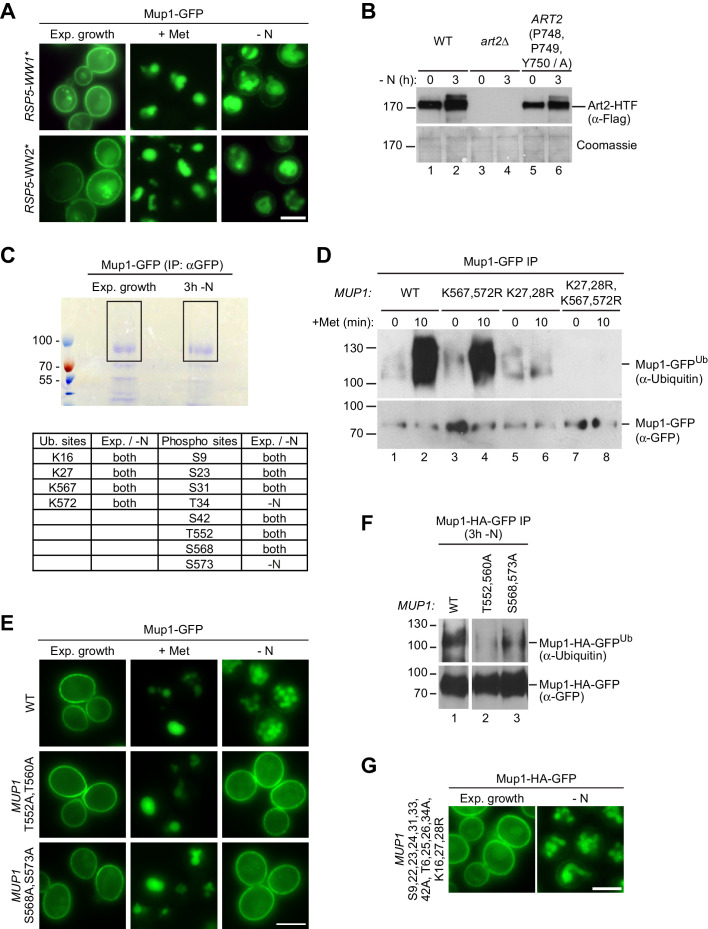
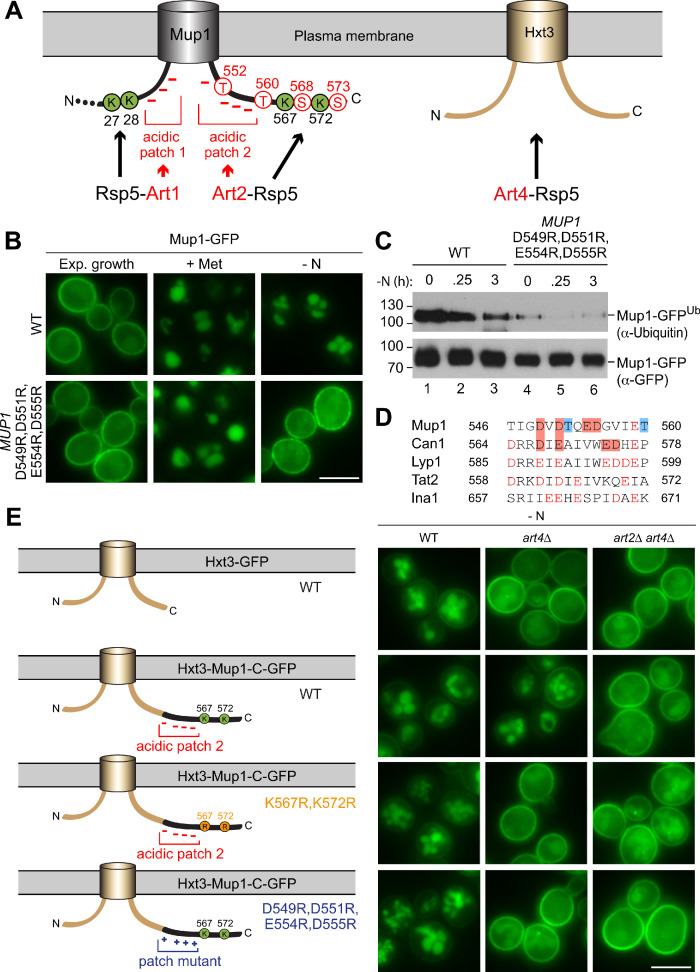
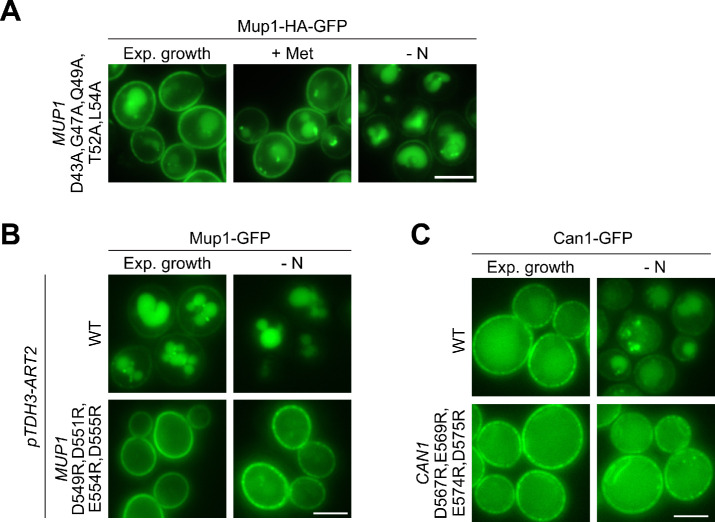
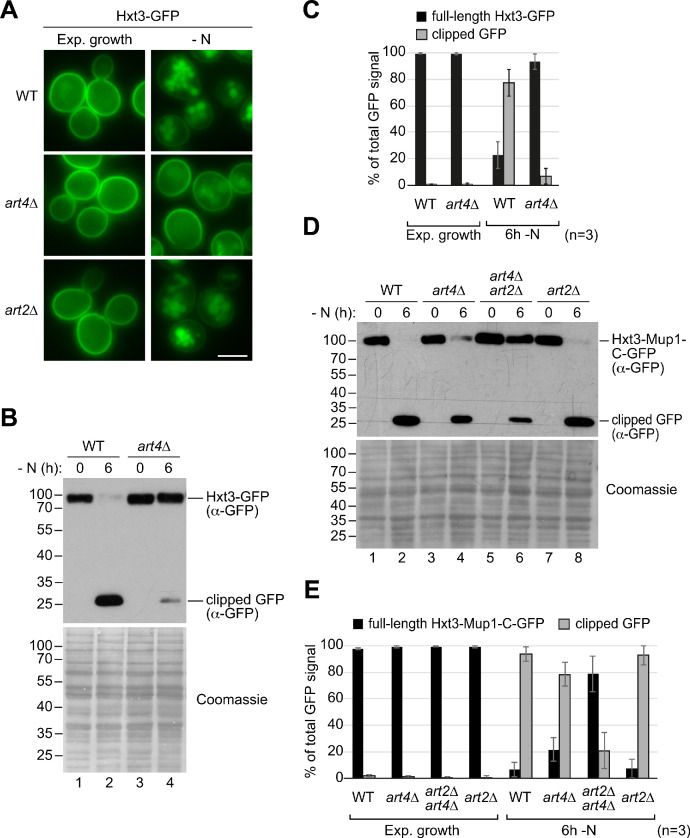
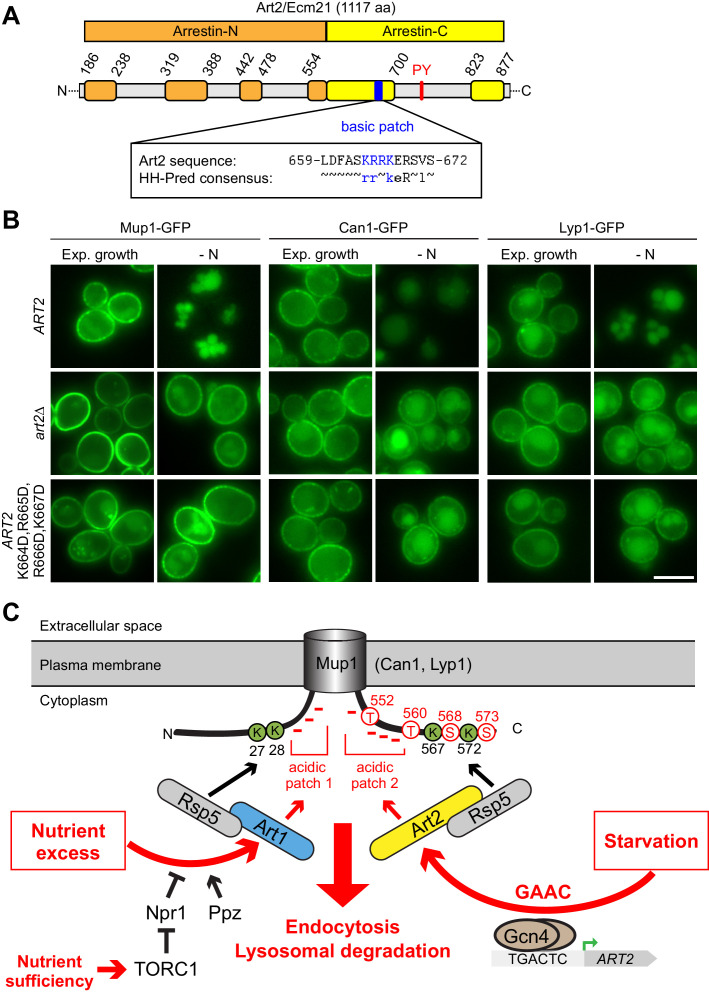
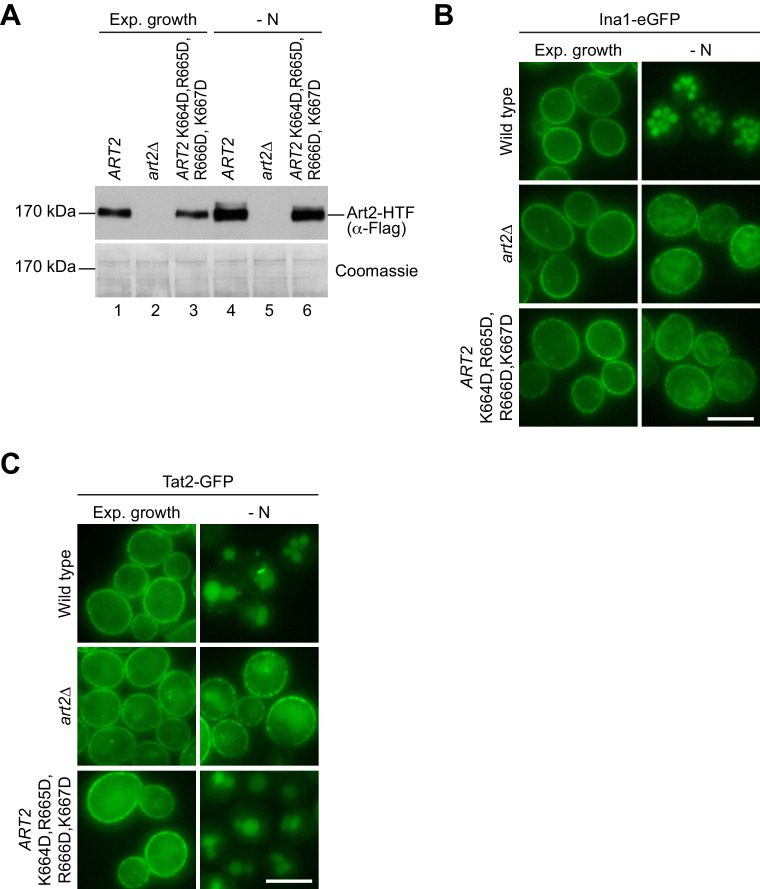
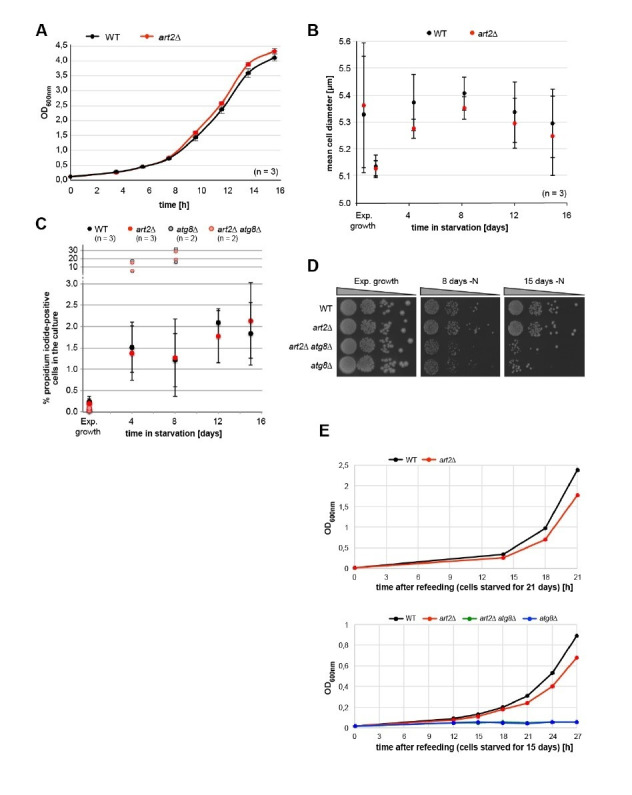
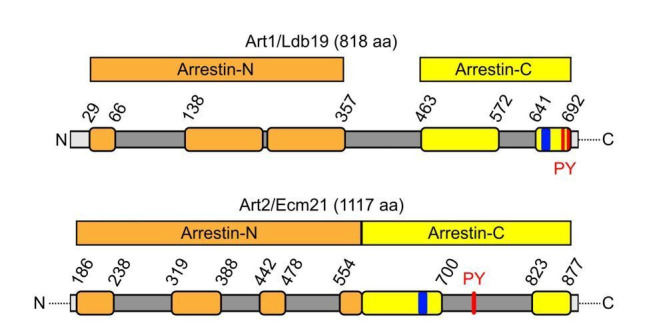
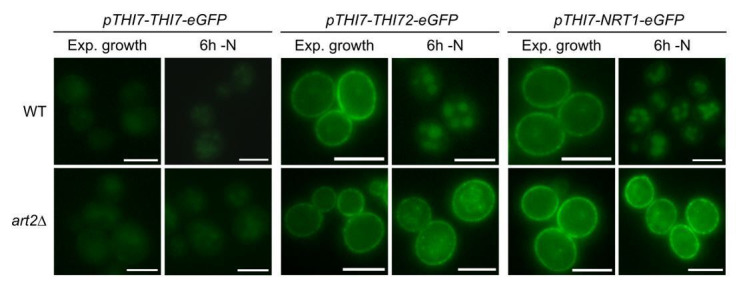
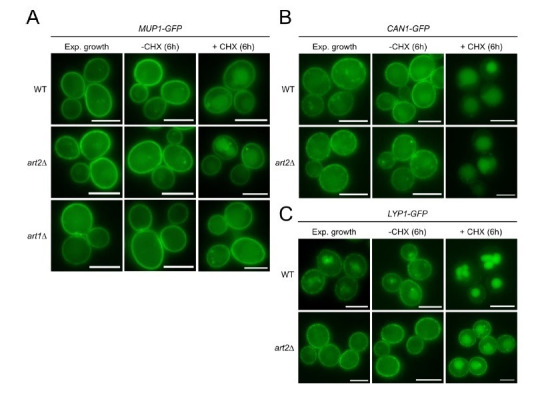
How cells adjust nutrient transport across their membranes is incompletely understood. Previously, we have shown that broadly re-configures the nutrient transporters at the plasma membrane in response to amino acid availability, through endocytosis of sugar- and amino acid transporters (AATs) (Müller et al., 2015). A genome-wide screen now revealed that the selective endocytosis of four AATs during starvation required the α-arrestin family protein Art2/Ecm21, an adaptor for the ubiquitin ligase Rsp5, and its induction through the general amino acid control pathway. Art2 uses a basic patch to recognize C-terminal acidic sorting motifs in AATs and thereby instructs Rsp5 to ubiquitinate proximal lysine residues. When amino acids are in excess, Rsp5 instead uses TORC1-activated Art1 to detect N-terminal acidic sorting motifs within the same AATs, which initiates exclusive substrate-induced endocytosis. Thus, amino acid excess or starvation activate complementary α-arrestin-Rsp5-complexes to control selective endocytosis and adapt nutrient acquisition.
细胞如何调节跨膜的营养物质运输还不完全清楚。以前,我们已经表明,广泛地重新配置了质膜上的营养物质转运体,以响应氨基酸的可用性,通过糖和氨基酸转运体(AATs)的内吞作用(Müller 等人,2015 年)。现在,全基因组筛选揭示了在饥饿期间选择性内吞作用的四种 AATs 需要α-抑制素家族蛋白 Art2/Ecm21,这是泛素连接酶 Rsp5 的衔接子,并且通过一般氨基酸控制途径诱导。Art2 使用碱性斑来识别 AATs 中的 C 末端酸性分拣基序,从而指示 Rsp5 使邻近的赖氨酸残基泛素化。当氨基酸过剩时,Rsp5 转而使用 TORC1 激活的 Art1 来检测同一 AATs 内的 N 末端酸性分拣基序,这会引发专有的底物诱导内吞作用。因此,氨基酸过剩或饥饿会激活互补的α-抑制素-Rsp5 复合物来控制选择性内吞作用和适应营养物质获取。