Center for Life Sciences, School of Life Sciences, Yunnan University, Kunming, Yunnan, China; Department of Hepatobiliary Surgery, The Second Affiliated Hospital of Guangdong Medical University, Zhanjiang, Guangdong, China; Department of Hepatobiliary Surgery, Affiliated Hospital of Guangdong Medical University, Zhanjiang, Guangdong, China; Key Laboratory of Animal Models and Human Disease Mechanisms of the Chinese Academy of Sciences and Yunnan Province, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, China.
Key Laboratory of Animal Models and Human Disease Mechanisms of the Chinese Academy of Sciences and Yunnan Province, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, China.
Cell Mol Gastroenterol Hepatol. 2021;11(3):697-724. doi: 10.1016/j.jcmgh.2020.10.006. Epub 2020 Oct 17.
BACKGROUND & AIMS: Nonalcoholic fatty liver disease (NAFLD) is becoming a severe liver disorder worldwide. Autophagy plays a critical role in liver steatosis. However, the role of autophagy in NAFLD remains exclusive and under debate. In this study, we investigated the role of S100 calcium binding protein A11 (S100A11) in the pathogenesis of hepatic steatosis.
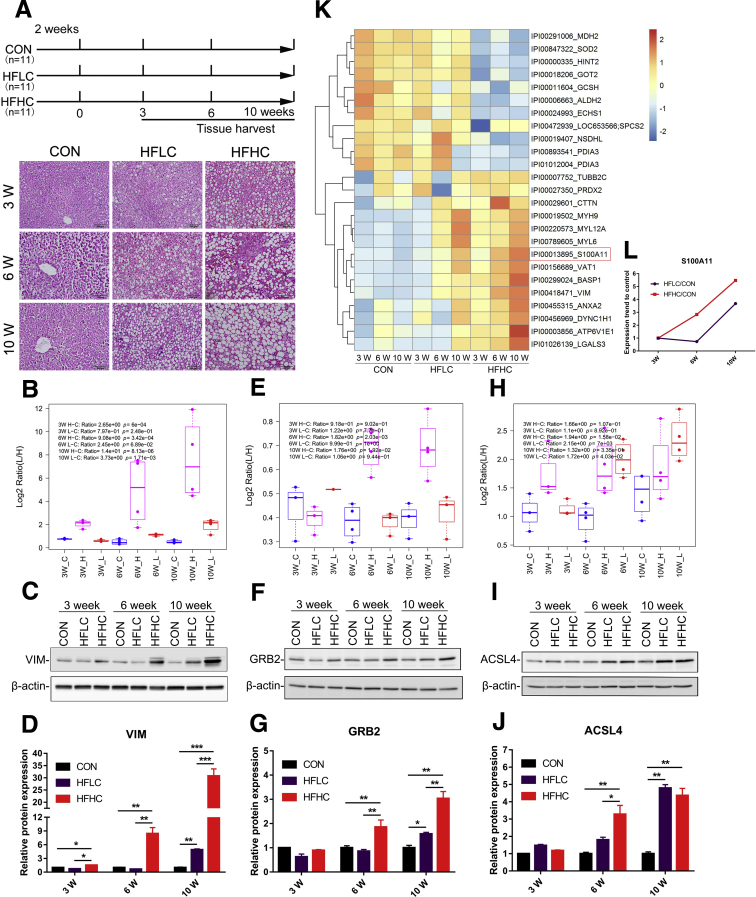
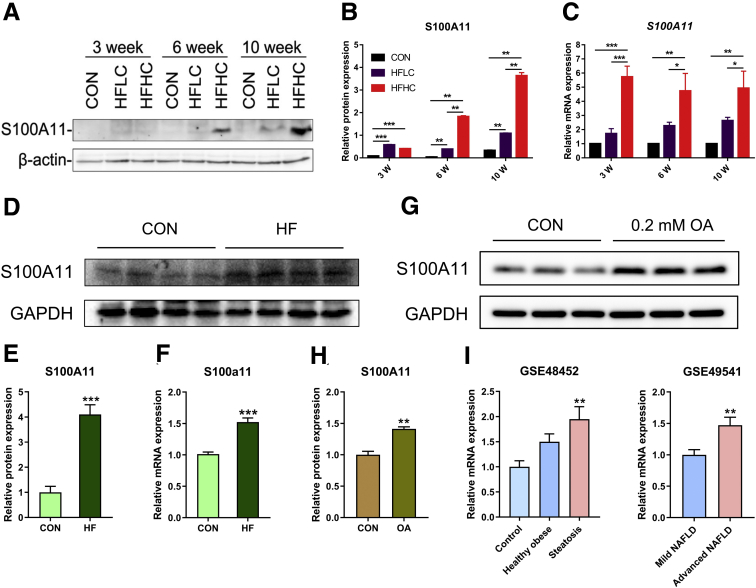
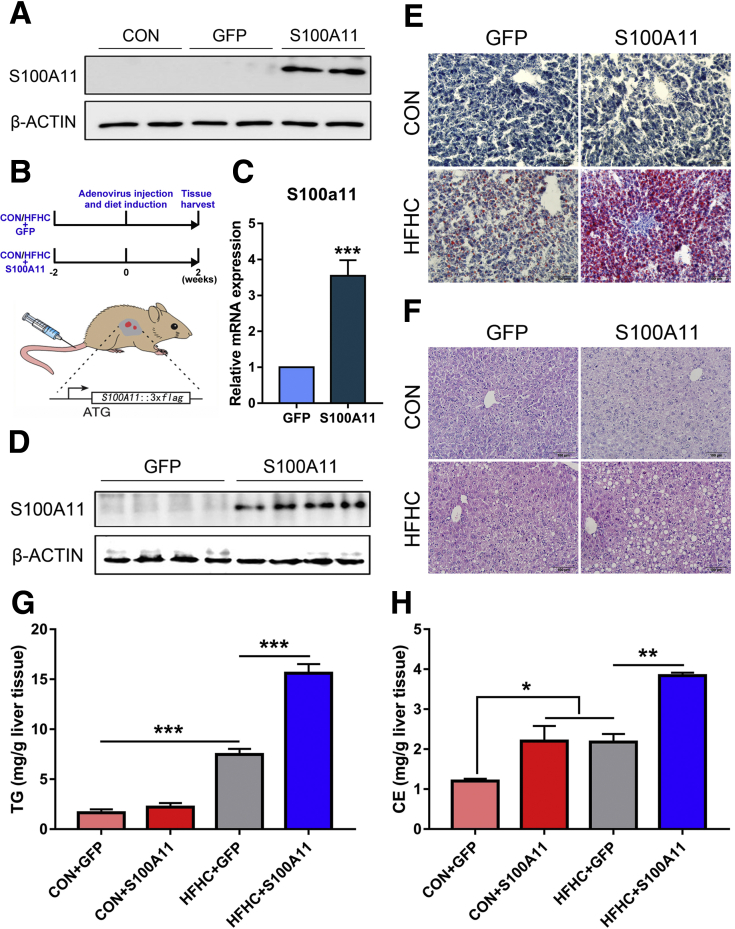
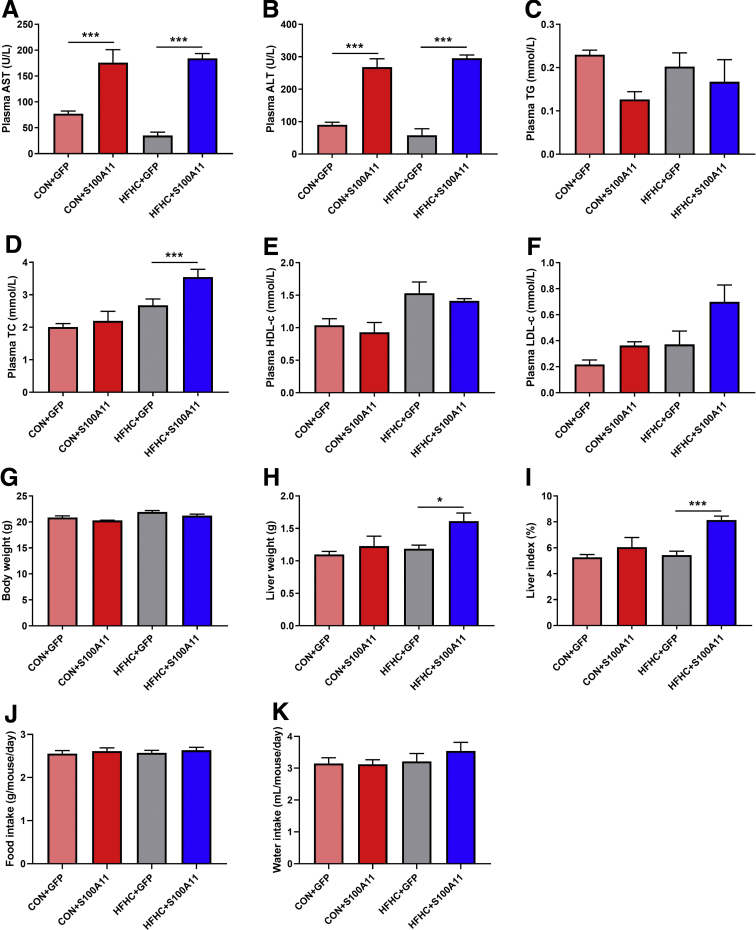
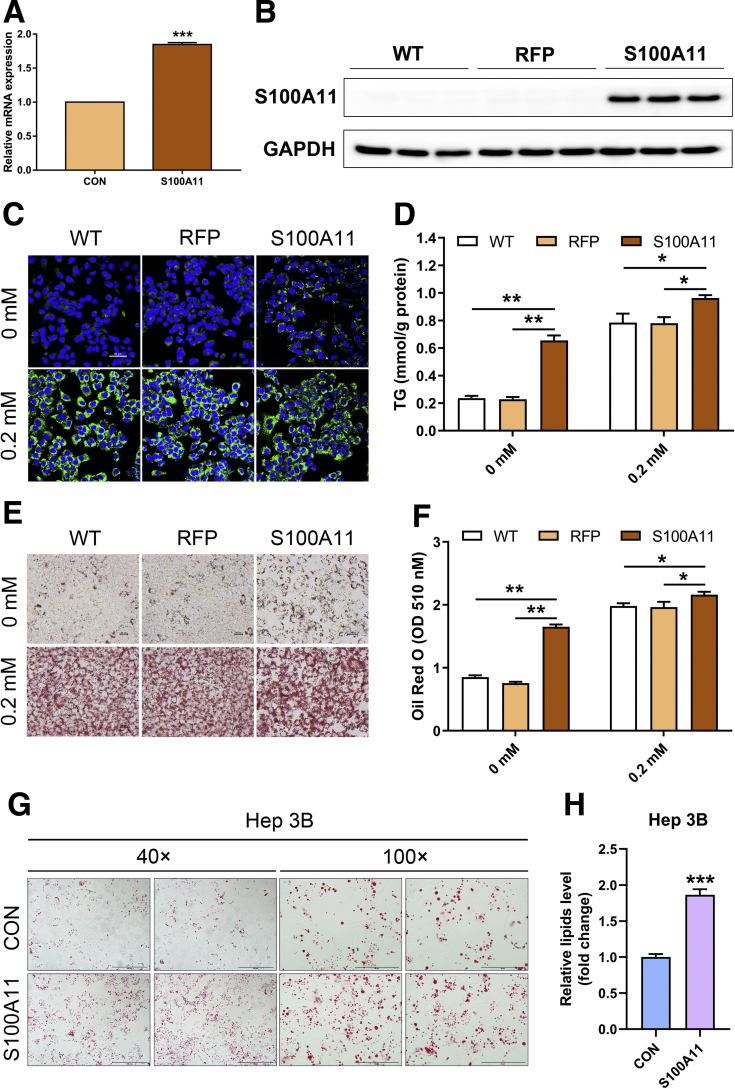
We performed liver proteomics in a well-established tree shrew model of NAFLD. The expression of S100A11 in different models of NAFLD was detected by Western blot and/or quantitative polymerase chain reaction. Liver S100A11 overexpression mice were generated by injecting a recombinant adenovirus gene transfer vector through the tail vein and then induced by a high-fat and high-cholesterol diet. Cell lines with S100a11 stable overexpression were established with a recombinant lentiviral vector. The lipid content was measured with either Bodipy staining, Oil Red O staining, gas chromatography, or a triglyceride kit. The autophagy and lipogenesis were detected in vitro and in vivo by Western blot and quantitative polymerase chain reaction. The functions of Sirtuin 1, histone deacetylase 6 (HDAC6), and FOXO1 were inhibited by specific inhibitors. The interactions between related proteins were analyzed by a co-immunoprecipitation assay and immunofluorescence analysis.
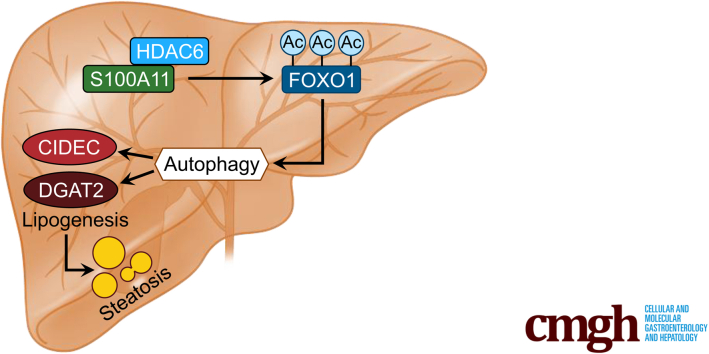
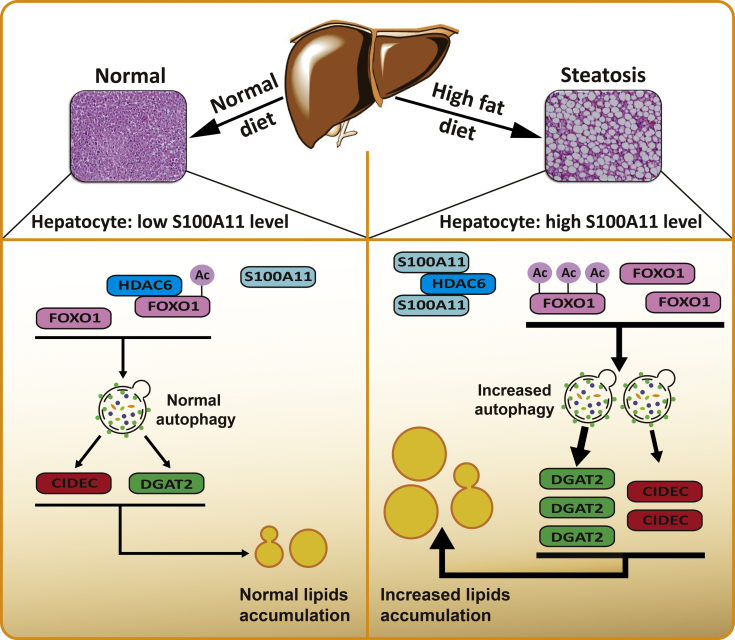
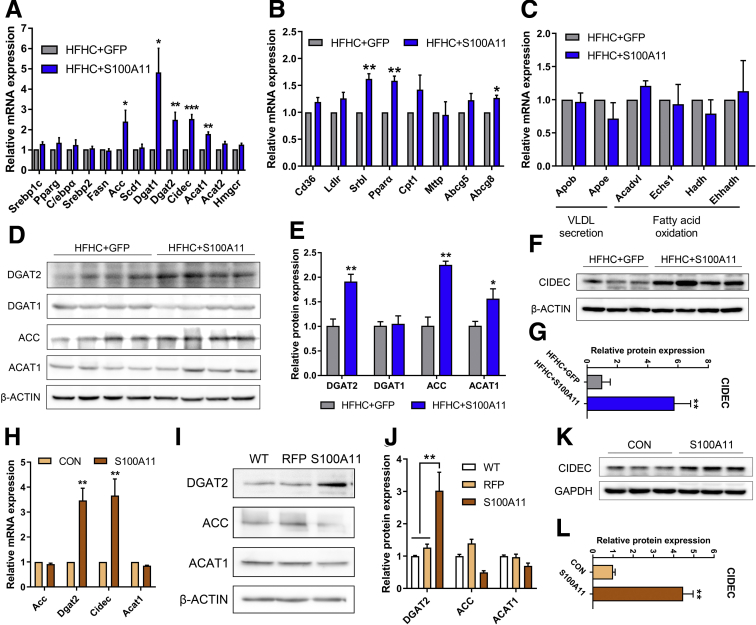
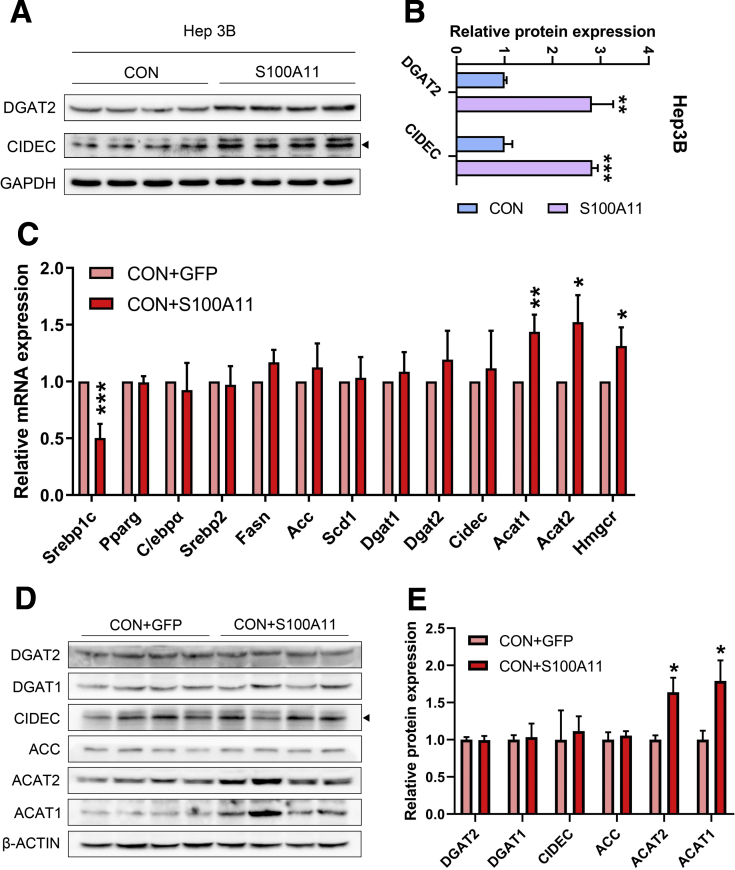
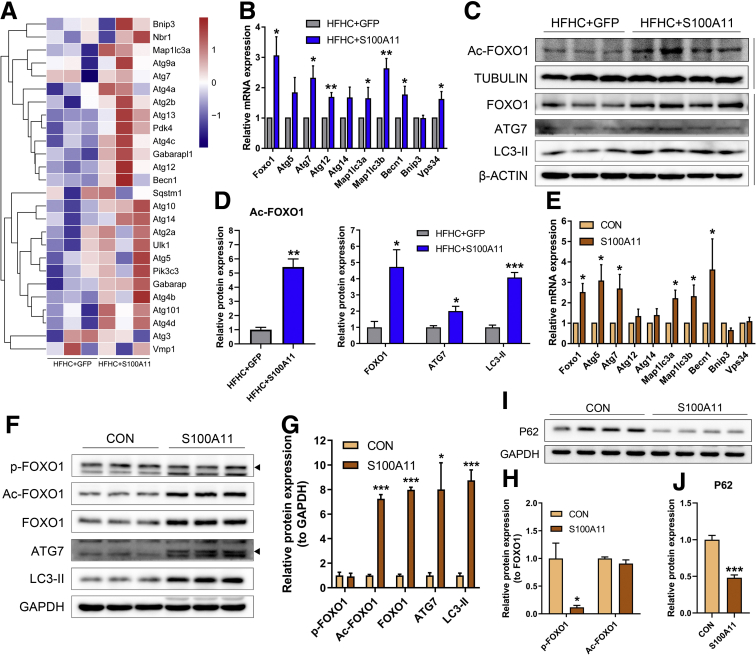
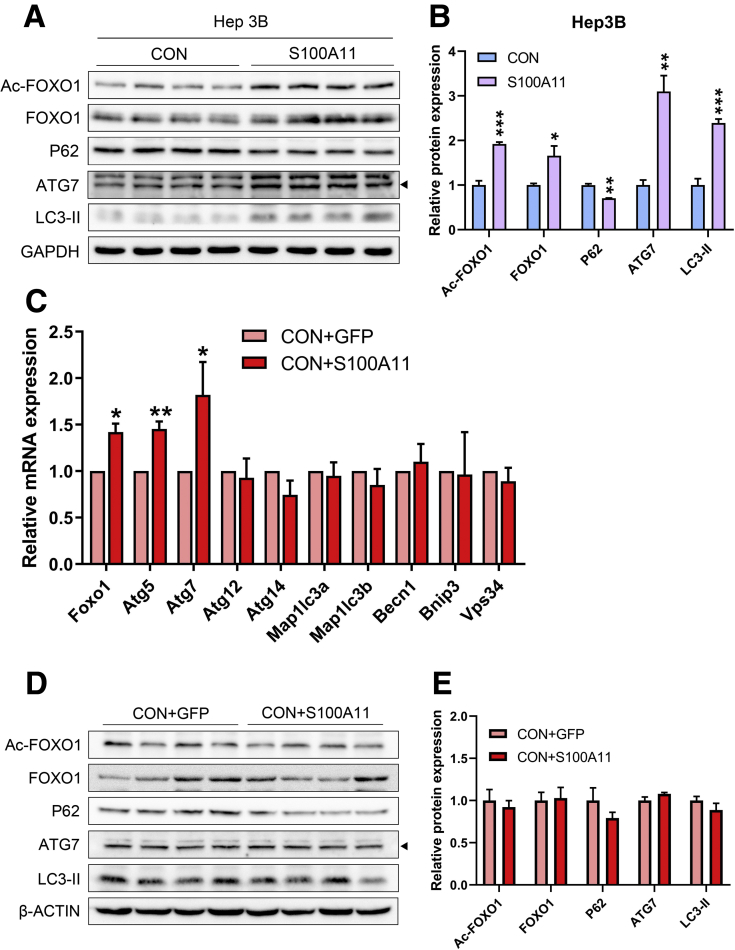
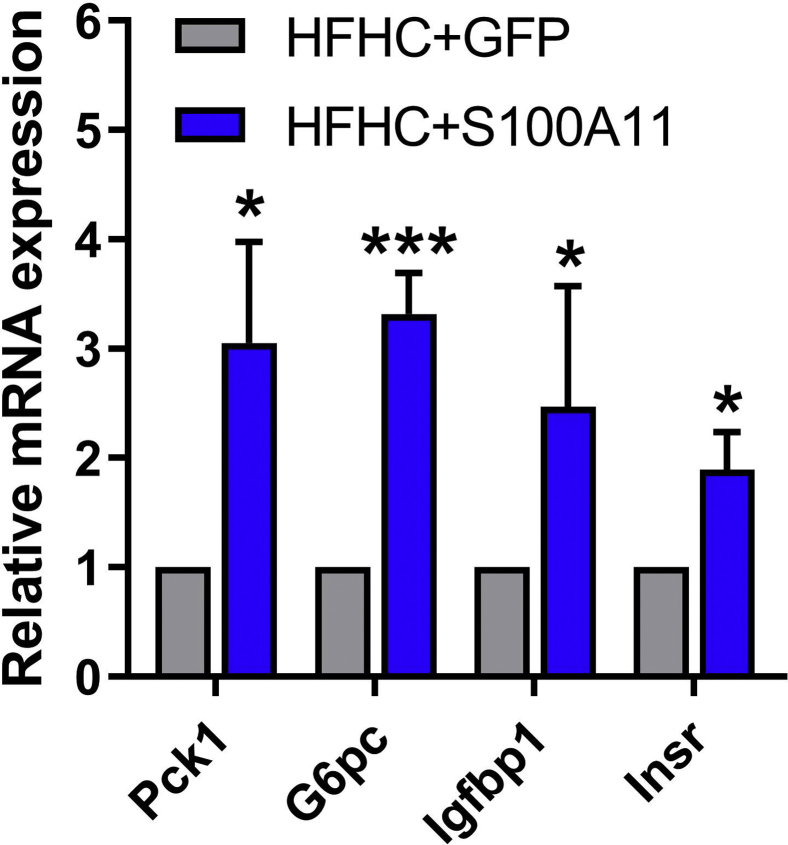
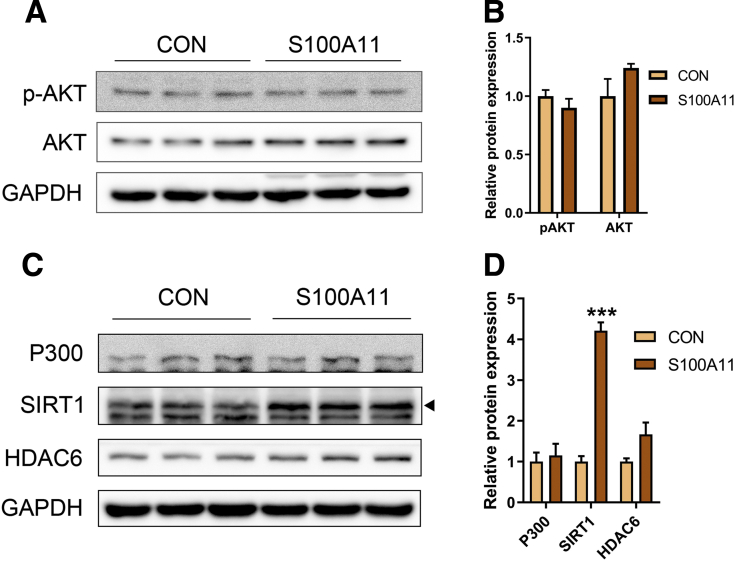
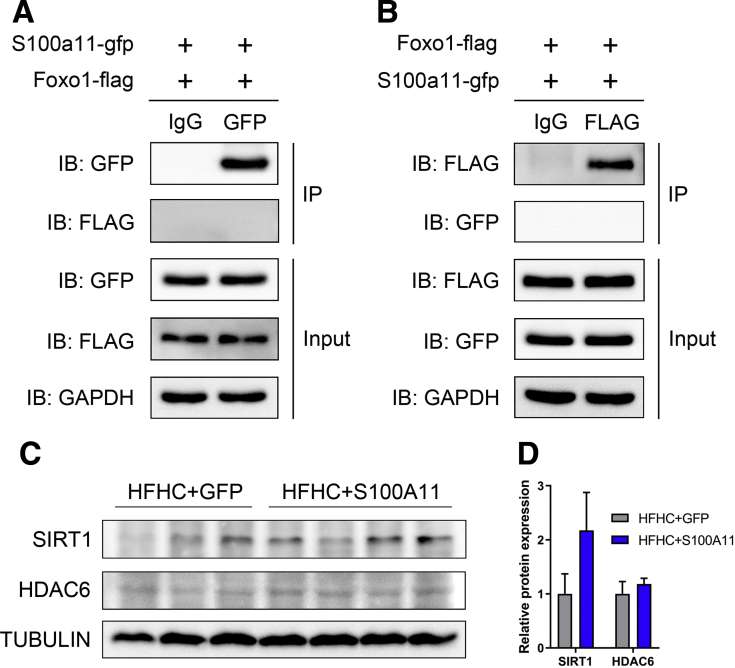
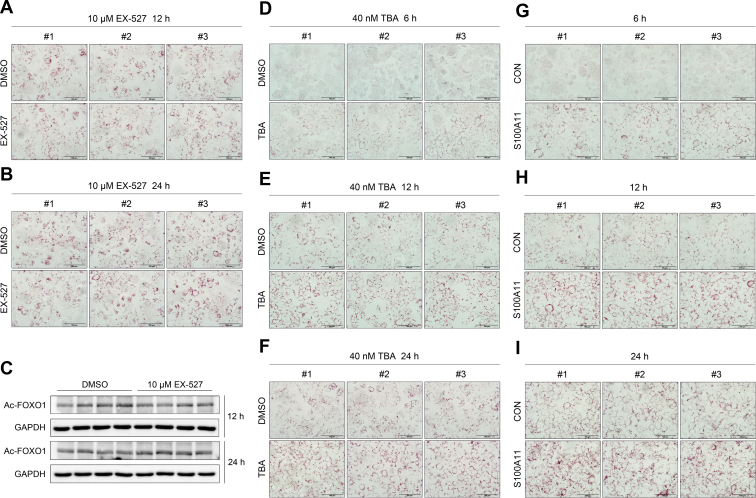
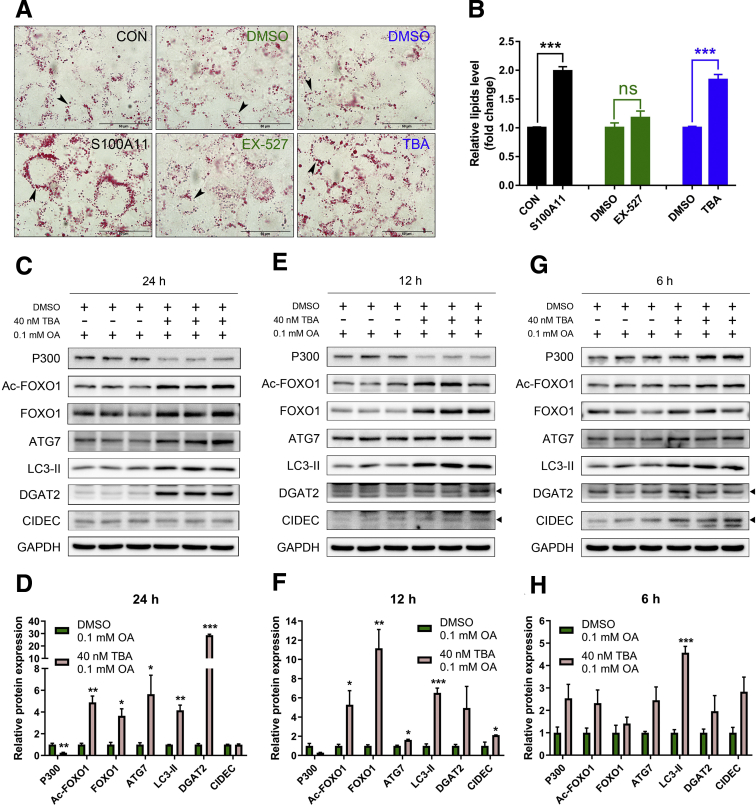
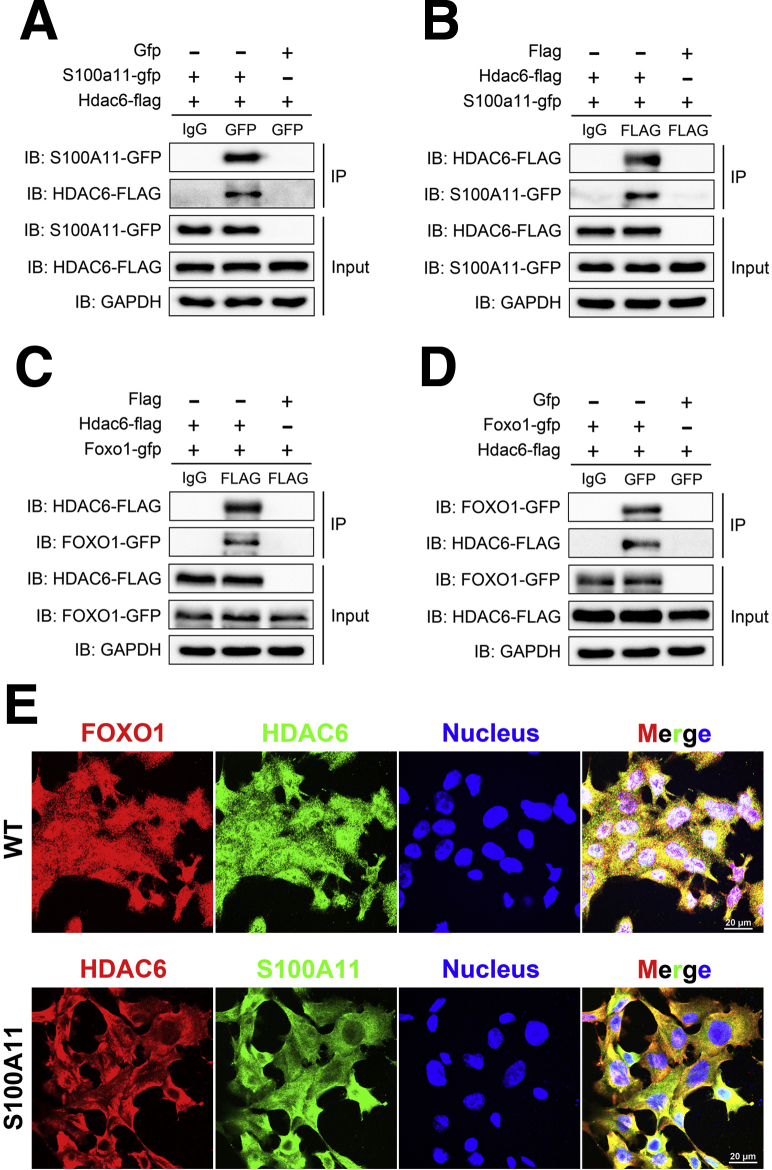
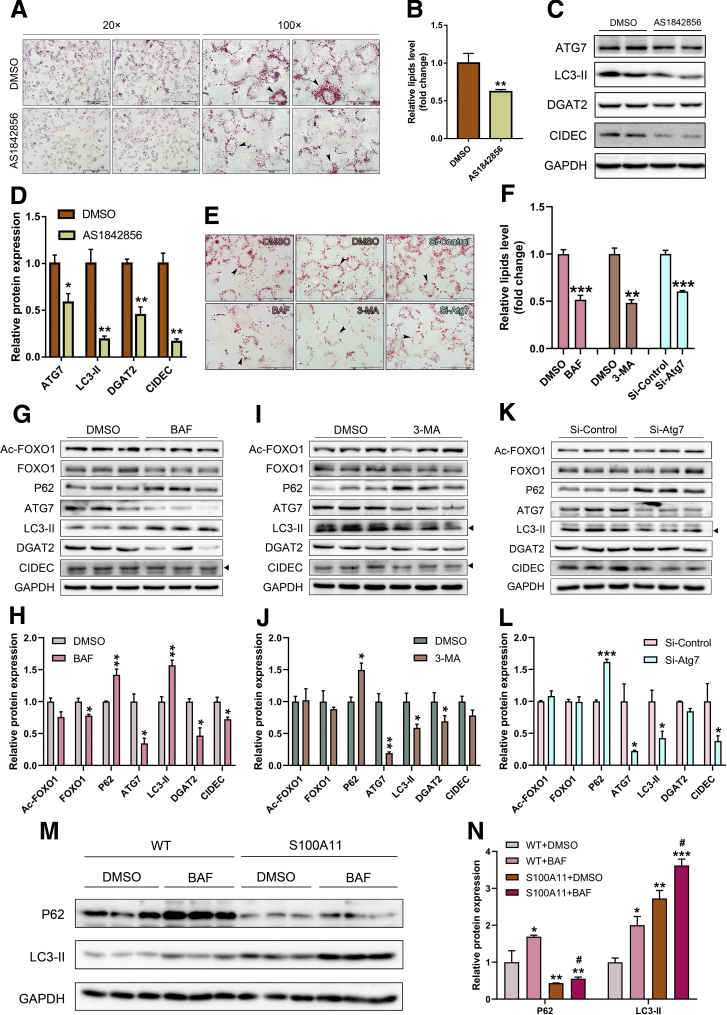
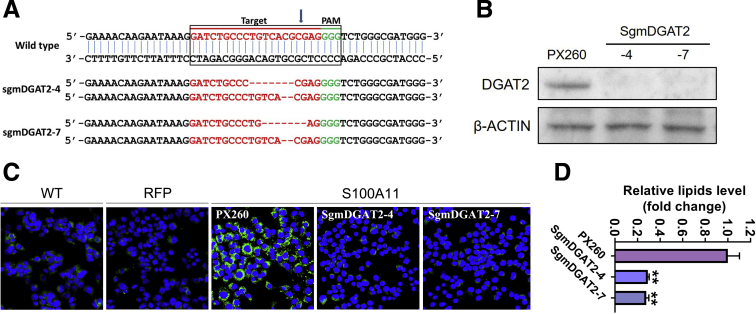
The expression of S100A11 was up-regulated significantly in a time-dependent manner in the tree shrew model of NAFLD. S100A11 expression was induced consistently in oleic acid-treated liver cells as well as the livers of mice fed a high-fat diet and NAFLD patients. Both in vitro and in vivo overexpression of S100A11 could induce hepatic lipid accumulation. Mechanistically, overexpression of S100A11 activated an autophagy and lipogenesis process through up-regulation and acetylation of the transcriptional factor FOXO1, consequently promoting lipogenesis and lipid accumulation in vitro and in vivo. Inhibition of HDAC6, a deacetylase of FOXO1, showed similar phenotypes to S100A11 overexpression in Hepa 1-6 cells. S100A11 interacted with HDAC6 to inhibit its activity, leading to the release and activation of FOXO1. Under S100A11 overexpression, the inhibition of FOXO1 and autophagy could alleviate the activated autophagy as well as up-regulated lipogenic genes. Both FOXO1 and autophagy inhibition and Dgat2 deletion could reduce liver cell lipid accumulation significantly.
A high-fat diet promotes liver S100A11 expression, which may interact with HDAC6 to block its binding to FOXO1, releasing or increasing the acetylation of FOXO1, thus activating autophagy and lipogenesis, and accelerating lipid accumulation and liver steatosis. These findings indicate a completely novel S100A11-HDAC6-FOXO1 axis in the regulation of autophagy and liver steatosis, providing potential possibilities for the treatment of NAFLD.
非酒精性脂肪性肝病(NAFLD)正在成为全球范围内一种严重的肝脏疾病。自噬在肝脂肪变性中起着关键作用。然而,自噬在 NAFLD 中的作用仍然是独特的,并且存在争议。在这项研究中,我们研究了 S100 钙结合蛋白 A11(S100A11)在肝脂肪变性发病机制中的作用。
我们在建立的树鼩 NAFLD 模型中进行了肝脏蛋白质组学研究。通过 Western blot 和/或定量聚合酶链反应检测不同 NAFLD 模型中 S100A11 的表达。通过尾静脉注射重组腺病毒基因转移载体生成肝脏 S100A11 过表达小鼠,然后用高脂肪高胆固醇饮食诱导。使用重组慢病毒载体建立 S100a11 稳定过表达的细胞系。通过 Bodipy 染色、油红 O 染色、气相色谱或甘油三酯试剂盒测量脂质含量。通过 Western blot 和定量聚合酶链反应在体外和体内检测自噬和脂生成。通过特异性抑制剂抑制 Sirtuin 1、组蛋白去乙酰化酶 6(HDAC6)和 FOXO1 的功能。通过共免疫沉淀分析和免疫荧光分析分析相关蛋白之间的相互作用。
S100A11 的表达在树鼩 NAFLD 模型中呈时间依赖性显著上调。油酸处理的肝细胞以及高脂肪饮食喂养的小鼠肝脏和 NAFLD 患者中 S100A11 的表达持续上调。S100A11 的体外和体内过表达均可诱导肝脂质堆积。机制上,S100A11 的过表达通过上调和乙酰化转录因子 FOXO1 激活自噬和脂生成过程,从而促进体外和体内的脂生成和脂质积累。在 Hepa 1-6 细胞中,抑制去乙酰化酶 HDAC6 可显示出与 S100A11 过表达相似的表型。S100A11 与 HDAC6 相互作用以抑制其活性,导致 FOXO1 的释放和激活。在 S100A11 过表达下,FOXO1 和自噬的抑制可减轻激活的自噬以及上调的脂生成基因。FOXO1 和自噬的抑制以及 Dgat2 缺失均可显著减少肝实质细胞脂质堆积。
高脂肪饮食促进肝脏 S100A11 的表达,可能与 HDAC6 相互作用,阻止其与 FOXO1 结合,释放或增加 FOXO1 的乙酰化,从而激活自噬和脂生成,加速脂质堆积和肝脂肪变性。这些发现表明 S100A11-HDAC6-FOXO1 轴在自噬和肝脂肪变性的调节中具有全新的作用,为 NAFLD 的治疗提供了潜在的可能性。