Departments of Biology, Massachusetts Institute of Technology, Cambridge, United States.
Chemistry, Massachusetts Institute of Technology, Cambridge, United States.
Elife. 2020 Oct 22;9:e61496. doi: 10.7554/eLife.61496.
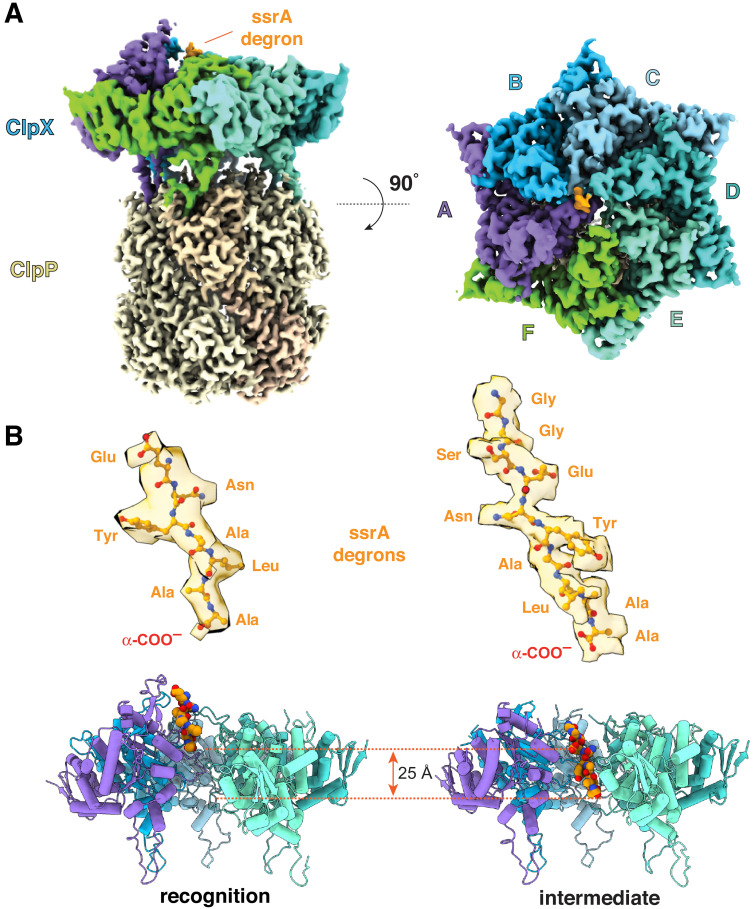
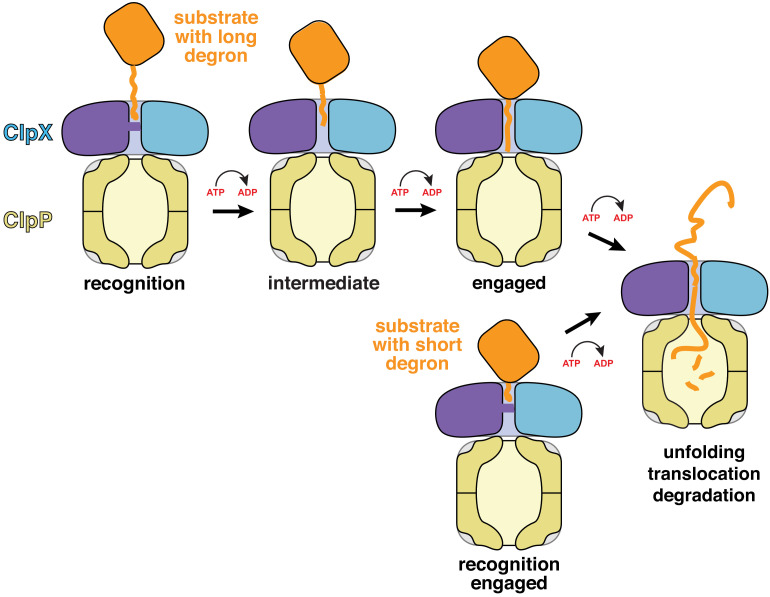
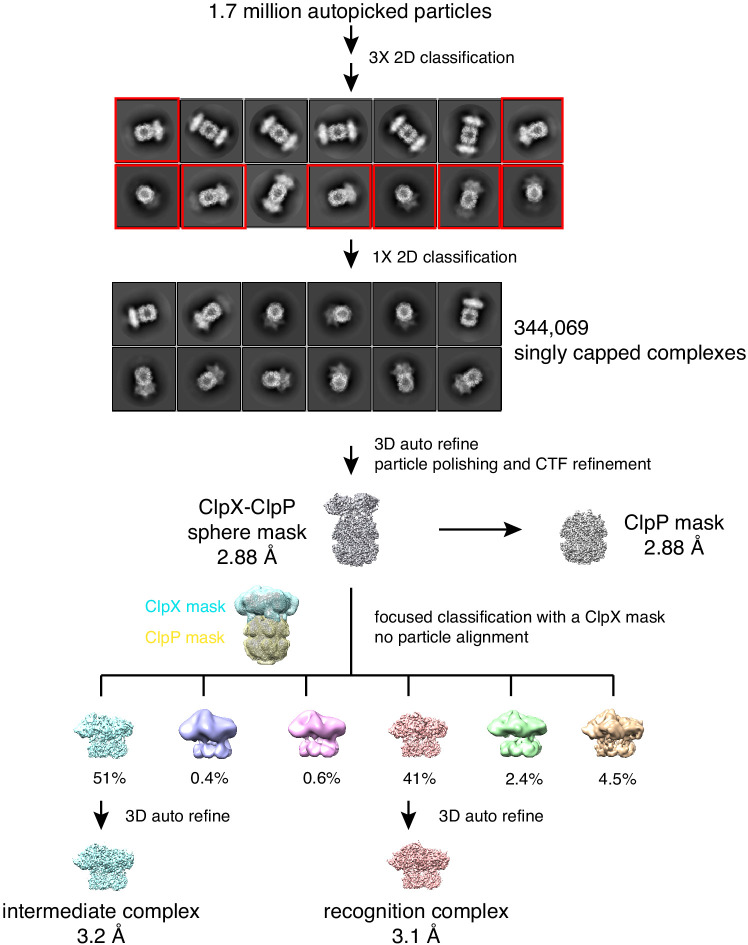
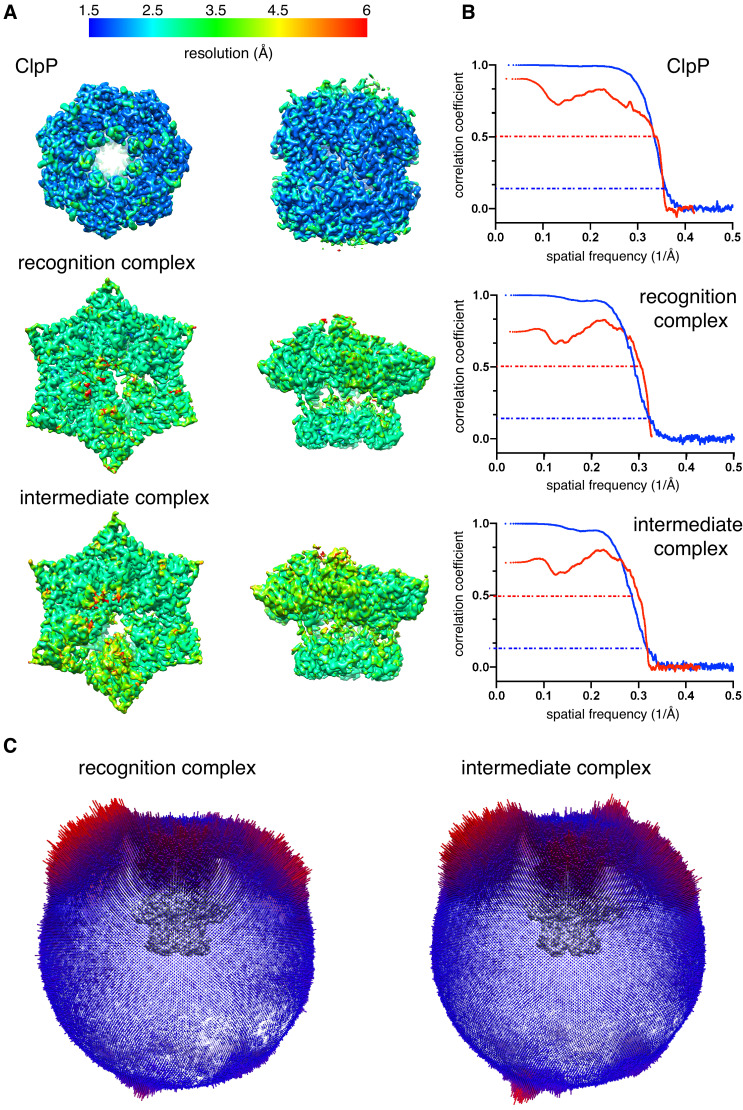
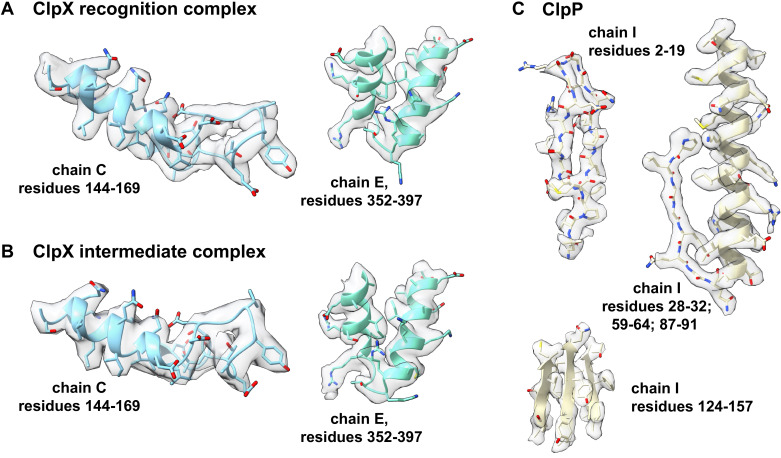
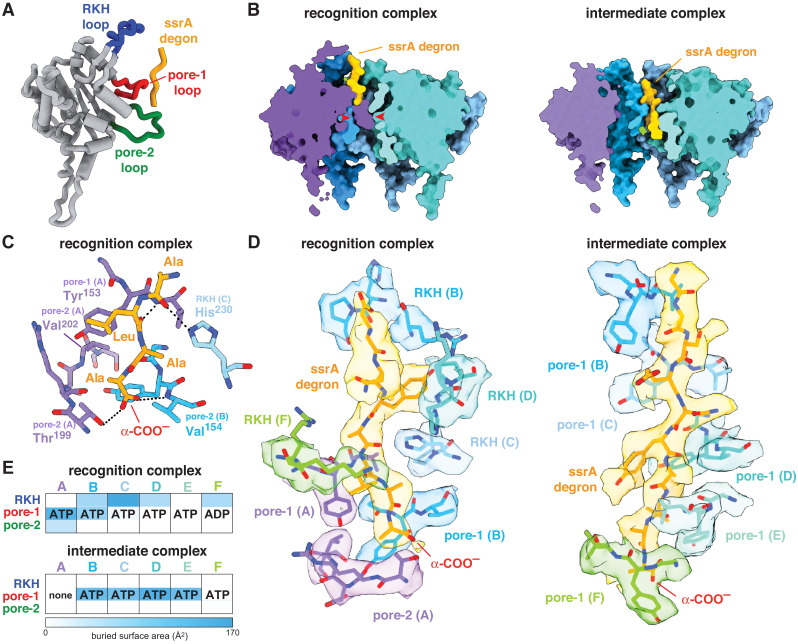
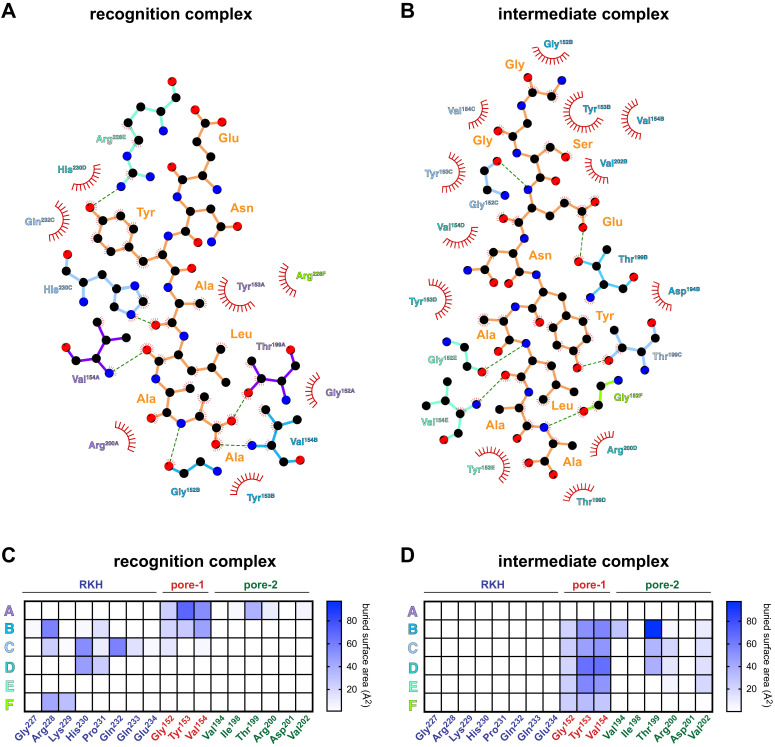
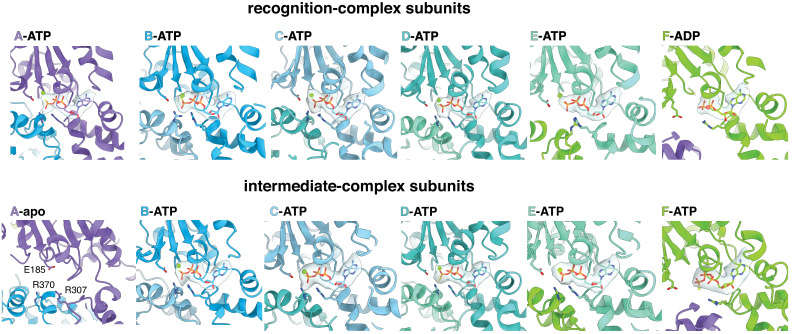
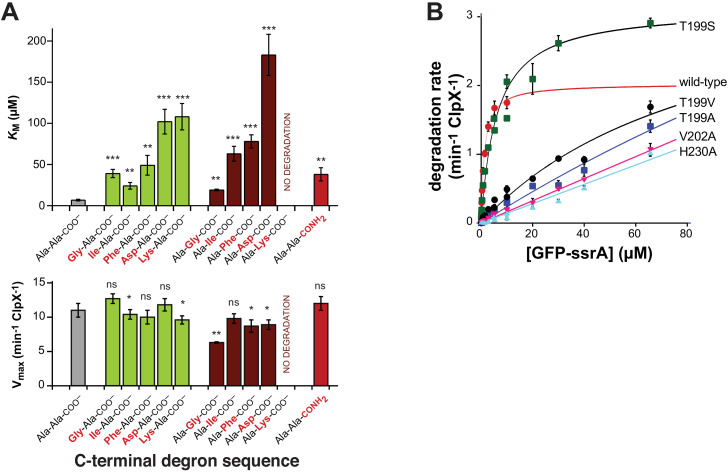
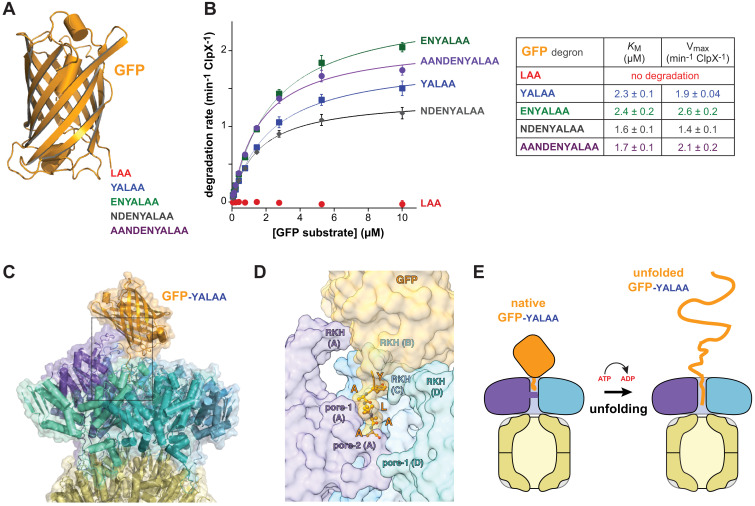
When ribosomes fail to complete normal translation, all cells have mechanisms to ensure degradation of the resulting partial proteins to safeguard proteome integrity. In and other eubacteria, the tmRNA system rescues stalled ribosomes and adds an ssrA tag or degron to the C-terminus of the incomplete protein, which directs degradation by the AAA+ ClpXP protease. Here, we present cryo-EM structures of ClpXP bound to the ssrA degron. C-terminal residues of the ssrA degron initially bind in the top of an otherwise closed ClpX axial channel and subsequently move deeper into an open channel. For short-degron protein substrates, we show that unfolding can occur directly from the initial closed-channel complex. For longer degron substrates, our studies illuminate how ClpXP transitions from specific recognition into a nonspecific unfolding and translocation machine. Many AAA+ proteases and protein-remodeling motors are likely to employ similar multistep recognition and engagement strategies.
当核糖体未能完成正常翻译时,所有细胞都有机制确保降解由此产生的部分蛋白质,以维护蛋白质组的完整性。在 和其他真细菌中,tmRNA 系统拯救了停滞的核糖体,并在不完全蛋白质的 C 末端添加一个 ssrA 标签或降解信号,指导 AAA+ ClpXP 蛋白酶进行降解。在这里,我们展示了 ClpXP 与 ssrA 降解信号结合的冷冻电镜结构。ssrA 降解信号的 C 末端残基最初结合在 ClpX 轴向通道的顶部,随后移动到更深处的开放通道。对于短降解信号的蛋白质底物,我们表明解折叠可以直接从初始的封闭通道复合物中发生。对于较长的降解信号底物,我们的研究阐明了 ClpXP 如何从特定识别转变为非特异性解折叠和易位机器。许多 AAA+ 蛋白酶和蛋白质重塑马达可能采用类似的多步骤识别和结合策略。