Department of Molecular Biology and Genetics, Aarhus University, 8000 Aarhus, Denmark.
Department of Biomedicine, Aarhus University, 8000 Aarhus, Denmark.
Nucleic Acids Res. 2020 Nov 18;48(20):11421-11433. doi: 10.1093/nar/gkaa873.
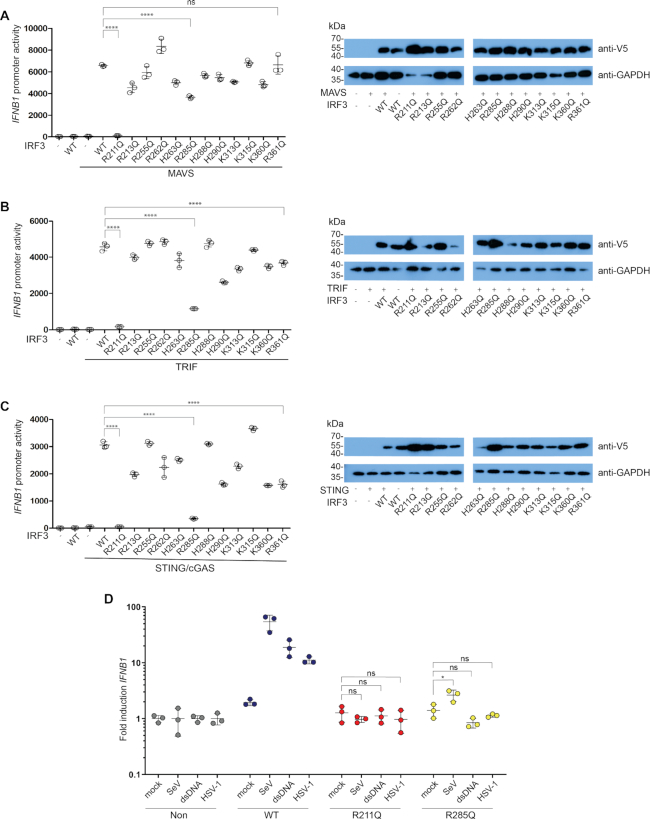
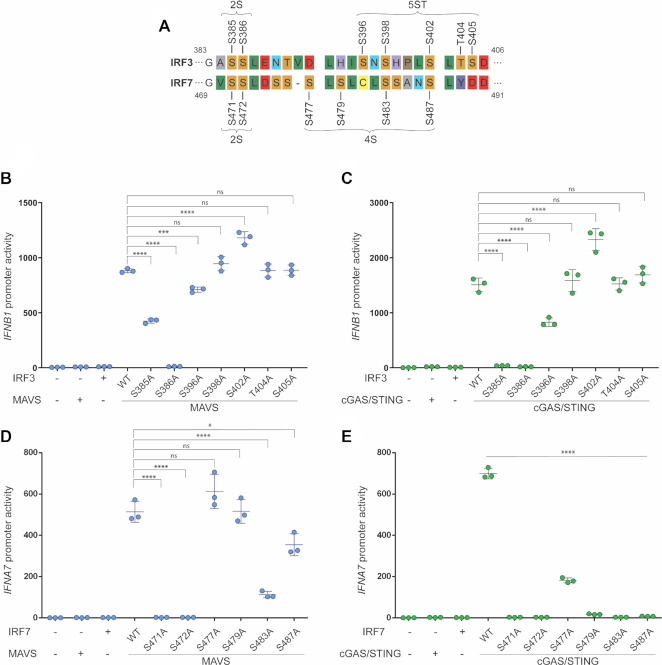
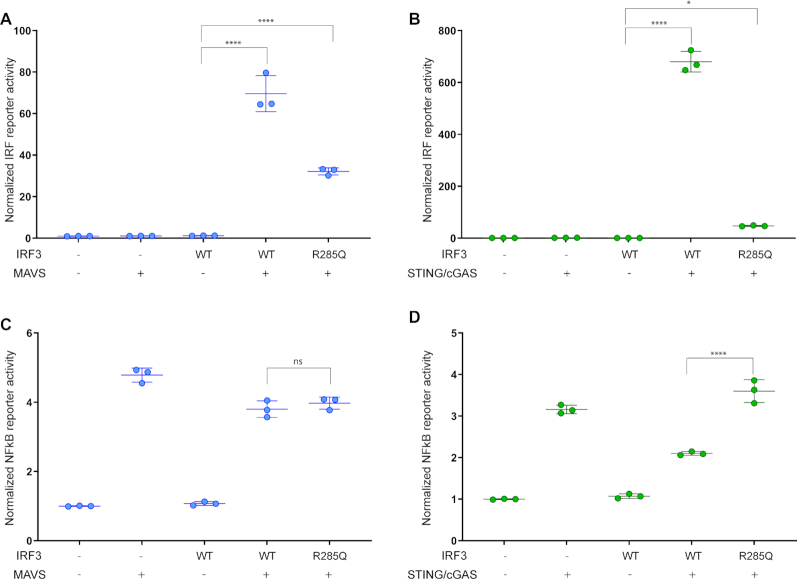
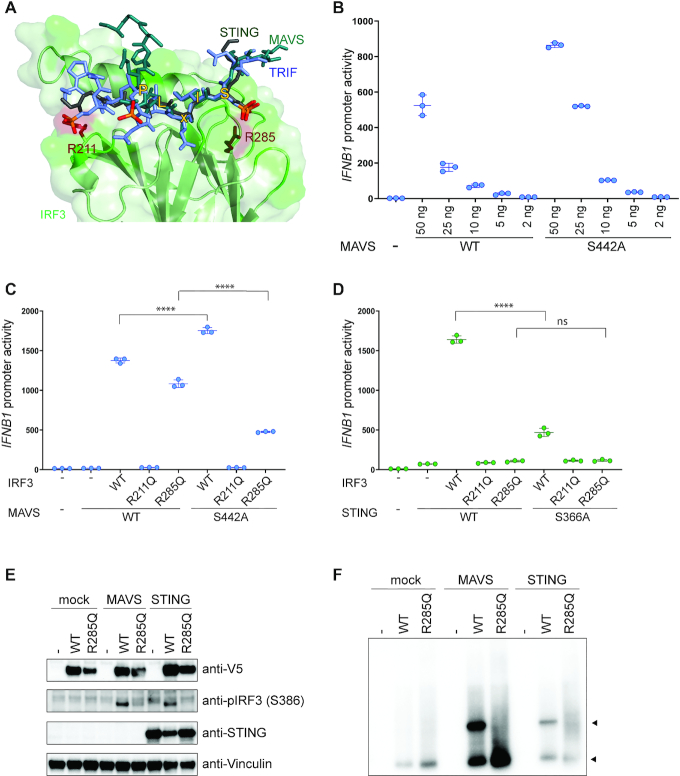
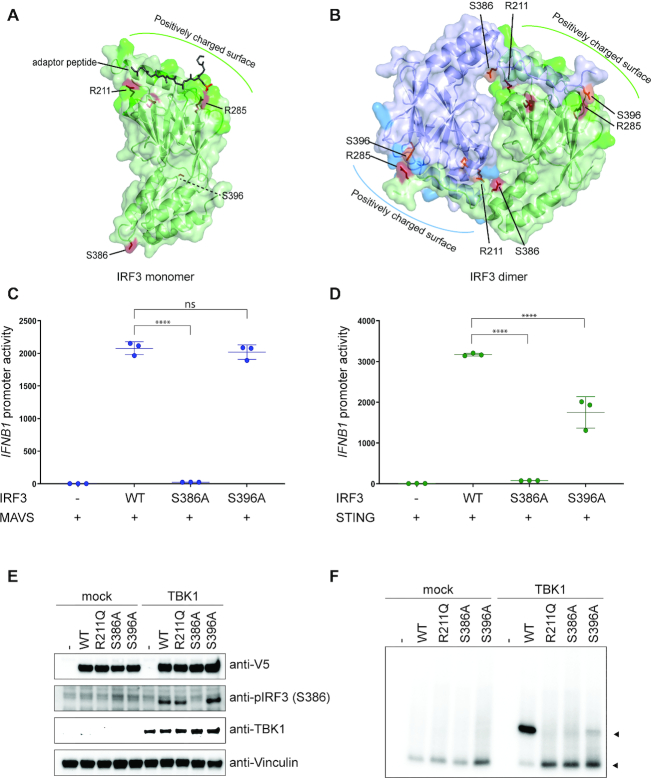
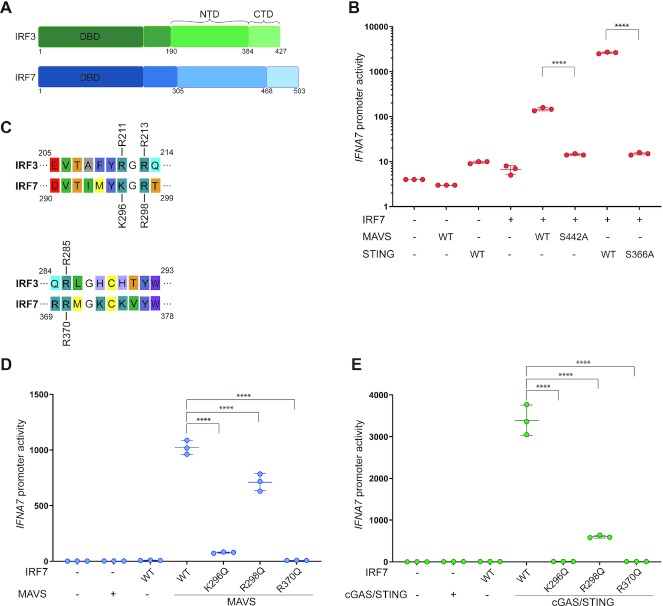
IRF3 and IRF7 are critical transcription factors in the innate immune response. Their activation is controlled by phosphorylation events, leading to the formation of homodimers that are transcriptionally active. Phosphorylation occurs when IRF3 is recruited to adaptor proteins via a positively charged surface within the regulatory domain of IRF3. This positively charged surface also plays a crucial role in forming the active homodimer by interacting with the phosphorylated sites stabilizing the homodimer. Here, we describe a distinct molecular interaction that is responsible for adaptor docking and hence phosphorylation as well as a separate interaction responsible for the formation of active homodimer. We then demonstrate that IRF7 can be activated by both MAVS and STING in a manner highly similar to that of IRF3 but with one key difference. Regulation of IRF7 appears more tightly controlled; while a single phosphorylation event is sufficient to activate IRF3, at least two phosphorylation events are required for IRF7 activation.
IRF3 和 IRF7 是先天免疫反应中的关键转录因子。它们的激活受磷酸化事件控制,导致形成转录活性的同源二聚体。当 IRF3 通过 IRF3 调节域内的正电荷表面被招募到衔接蛋白时,就会发生磷酸化。这个正电荷表面在通过与稳定同源二聚体的磷酸化位点相互作用形成活性同源二聚体方面也起着至关重要的作用。在这里,我们描述了一个负责衔接蛋白 docking 从而磷酸化的独特分子相互作用,以及一个负责形成活性同源二聚体的单独相互作用。然后,我们证明 IRF7 可以通过 MAVS 和 STING 以与 IRF3 非常相似的方式被激活,但有一个关键区别。IRF7 的调节似乎受到更严格的控制;虽然单个磷酸化事件足以激活 IRF3,但至少需要两个磷酸化事件才能激活 IRF7。