Key Laboratory of Molecular Biophysics of Ministry of Education, School of Life Science & Technology, Huazhong University of Science & Technology, Wuhan, Hubei, China.
J Physiol. 2021 Feb;599(4):1151-1172. doi: 10.1113/JP280578. Epub 2020 Dec 9.
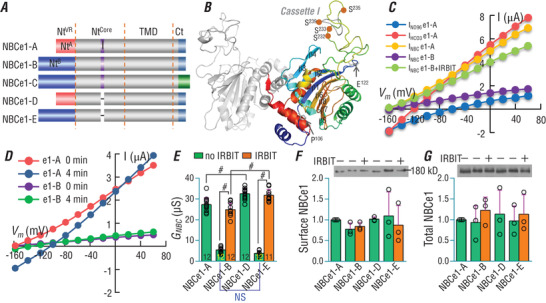
The electrogenic Na /HCO cotransporter NBCe1-B is widely expressed in many tissues, including pancreas, submandibular gland, brain, heart, etc. NBCe1-B has very low activity under basal condition due to auto-inhibition, but can be fully activated by protein interaction with the IP3R-binding protein released with inositol 1,4,5-trisphosphate (IRBIT). The structural components of the auto-inhibition domain and the IRBIT-binding domain of NBCe1-B are finely characterized based on systematic mutations in the present study and data from previous studies. Reducing negative charges on the cytosol side of the transmembrane domain greatly decreases the magnitude of the auto-inhibition of NBCe1-B. We propose that the auto-inhibition domain functions as a brake module that inactivates NBCe1-B by binding to, via electrostatic attraction, the transmembrane domain; IRBIT activates NBCe1-B by releasing the brake from the transmembrane domain via competitive binding to the auto-inhibition domain.
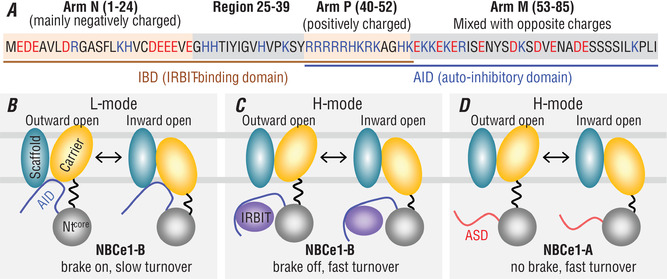
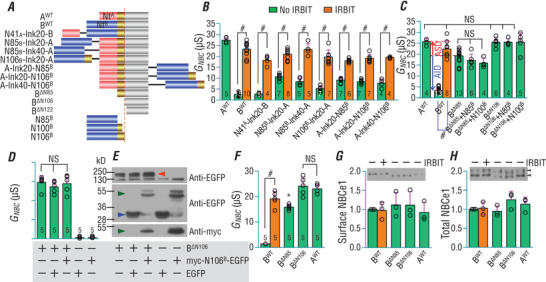
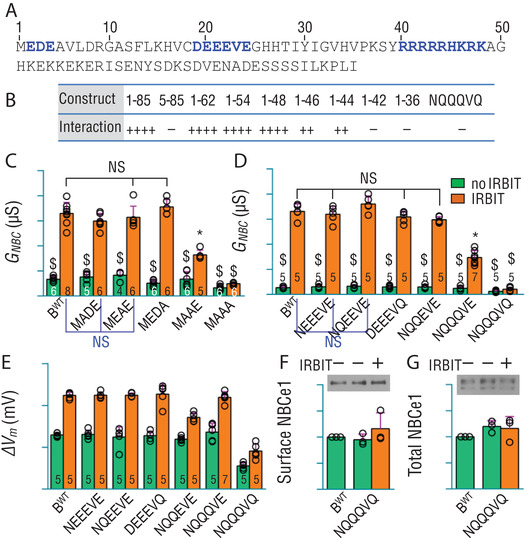
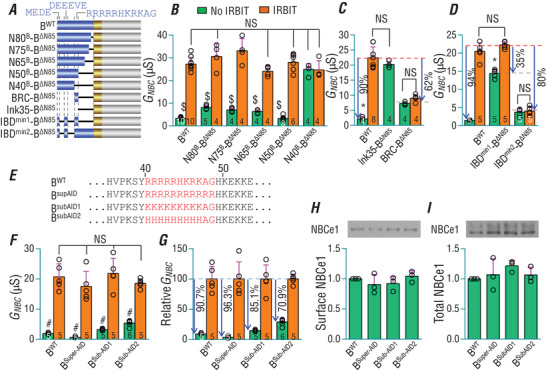
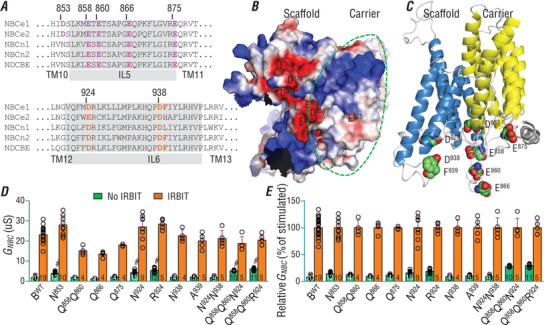
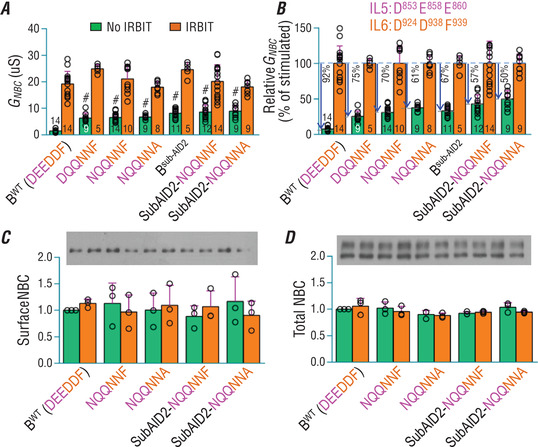
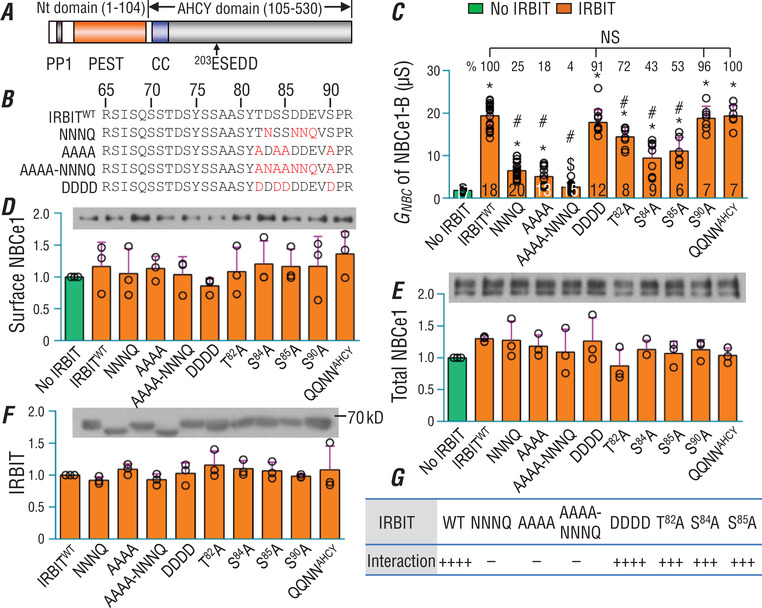
The electrogenic Na /HCO cotransporter NBCe1-B is widely expressed in many tissues in the body. NBCe1-B exhibits only basal activity due to the action of the auto-inhibition domain (AID) in its unique amino-terminus. However, NBCe1-B can be activated by interaction with the IP3R-binding protein released with inositol 1,4,5-trisphosphate (IRBIT). Here, we investigate the molecular mechanism underlying the auto-inhibition of NBCe1-B and its activation by IRBIT. The IRBIT-binding domain (IBD) of NBCe1-B spans residues 1-52, essentially consisting of two arms, one negatively charged (residues 1-24) and the other positively charged (residues 40-52). The AID mainly spans residues 40-85, overlapping with the IBD in the positively charged arm. The magnitude of auto-inhibition of NBCe1-B is greatly decreased by manipulating the positively charged residues in the AID or by replacing a set of negatively charged residues with neutral ones in the transmembrane domain. The interaction between IRBIT and NBCe1-B is abolished by mutating a set of negatively charged Asp/Glu residues (to Asn/Gln) plus a set of Ser/Thr residues (to Ala) in the PEST domain of IRBIT. However, this interaction is not affected by replacing the same set of Ser/Thr residues in the PEST domain with Asp. We propose that: (1) the AID, acting as a brake, binds to the transmembrane domain via electrostatic interaction to slow down NBCe1-B; (2) IRBIT activates NBCe1-B by releasing the brake from the transmembrane domain.
电中性的 Na+/HCO3-共转运蛋白 NBCe1-B 在许多组织中广泛表达,包括胰腺、颌下腺、大脑、心脏等。NBCe1-B 由于自身抑制作用,在基础条件下活性非常低,但可以通过与三磷酸肌醇(IP3)受体结合蛋白(IRBIT)结合释放而被完全激活。本研究通过系统突变和以前的研究数据,对 NBCe1-B 的自身抑制域和 IRBIT 结合域的结构组成进行了精细表征。跨膜域胞质侧负电荷的减少大大降低了 NBCe1-B 的自身抑制程度。我们提出,自身抑制域作为一个制动模块,通过静电吸引与跨膜域结合,从而使 NBCe1-B 失活;IRBIT 通过与自身抑制域竞争性结合,从跨膜域释放制动,从而激活 NBCe1-B。
电中性的 Na+/HCO3-共转运蛋白 NBCe1-B 在体内许多组织中广泛表达。由于其独特的氨基末端的自身抑制域(AID)的作用,NBCe1-B 仅表现出基础活性。然而,NBCe1-B 可以通过与三磷酸肌醇(IP3)受体结合蛋白(IRBIT)的相互作用而被激活。在这里,我们研究了 NBCe1-B 自身抑制及其被 IRBIT 激活的分子机制。NBCe1-B 的 IRBIT 结合域(IBD)跨越残基 1-52,基本上由两个臂组成,一个带负电荷(残基 1-24),另一个带正电荷(残基 40-52)。AID 主要跨越残基 40-85,与正电荷臂中的 IBD 重叠。通过操纵 AID 中的正电荷残基或用跨膜域中的中性残基取代一组负电荷残基,NBCe1-B 的自身抑制程度大大降低。在 IRBIT 的 PEST 结构域中突变一组带负电荷的天冬氨酸/谷氨酸残基(突变为天冬酰胺/谷氨酰胺)和一组丝氨酸/苏氨酸残基(突变为丙氨酸),会使 IRBIT 与 NBCe1-B 的相互作用丧失。然而,这种相互作用不受用 PEST 结构域中的相同组丝氨酸/苏氨酸残基替换为天冬氨酸的影响。我们提出:(1)AID 作为一个刹车,通过静电相互作用与跨膜域结合,从而减缓 NBCe1-B 的速度;(2)IRBIT 通过从跨膜域释放刹车来激活 NBCe1-B。