Molecular Systems Group, Quantitative Biology Section, Medical Research Council London Institute of Medical Sciences, London W12 0NN, United Kingdom.
Institute of Clinical Sciences, Faculty of Medicine, Imperial College London, London W12 0NN, United Kingdom.
Proc Natl Acad Sci U S A. 2020 Dec 29;117(52):33384-33395. doi: 10.1073/pnas.2007056117. Epub 2020 Dec 7.
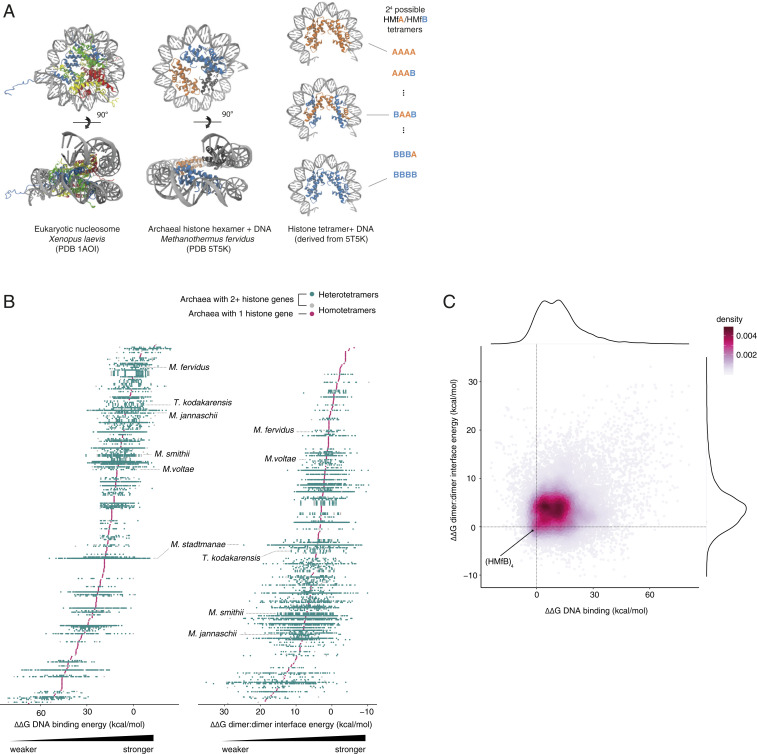
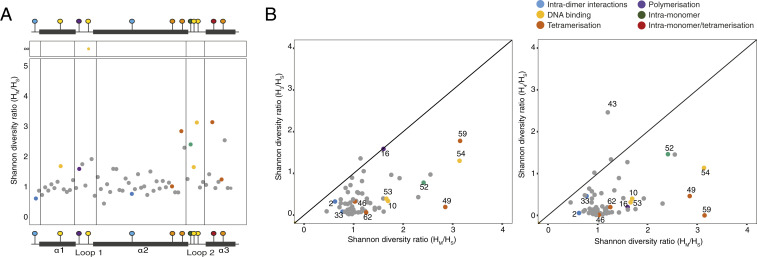
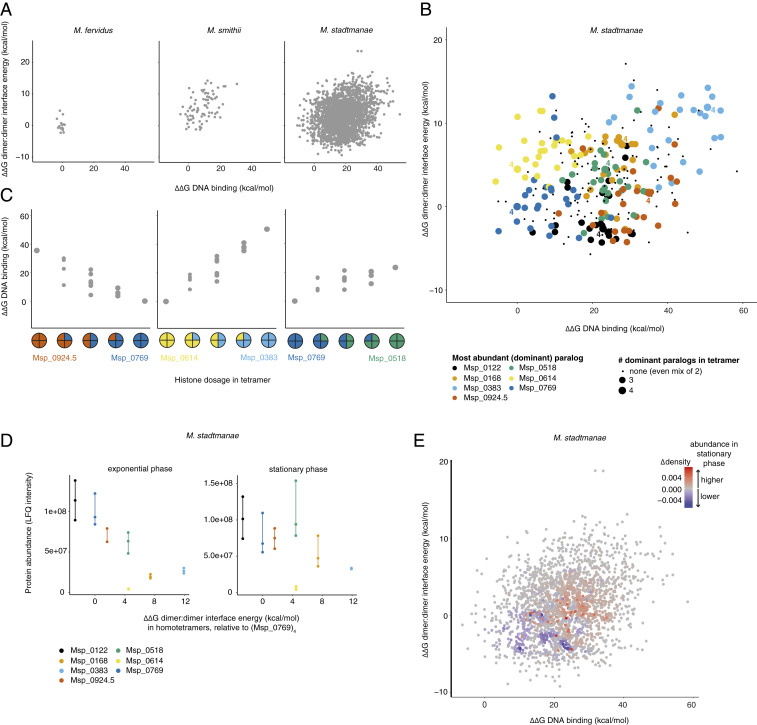
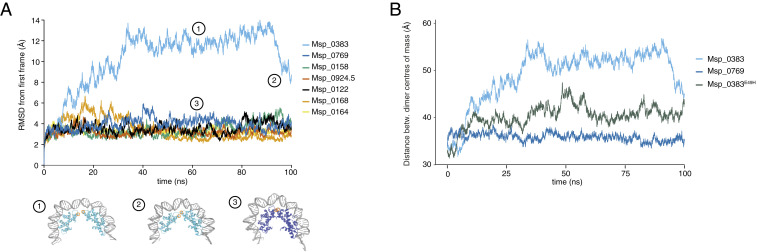
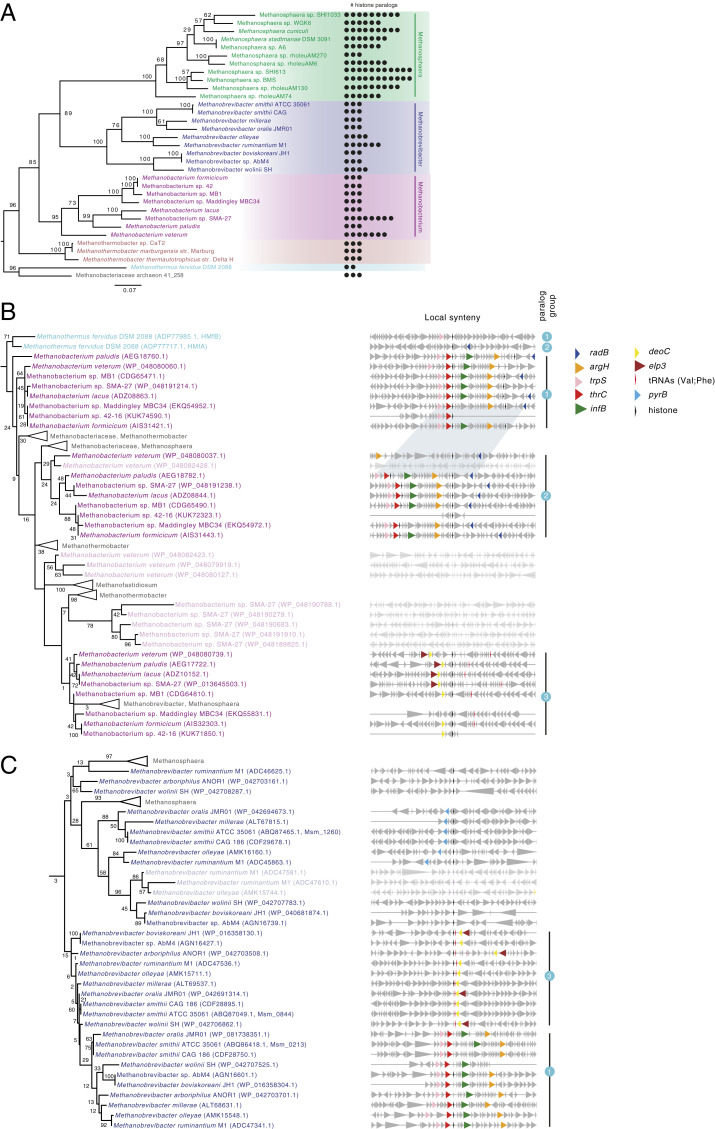
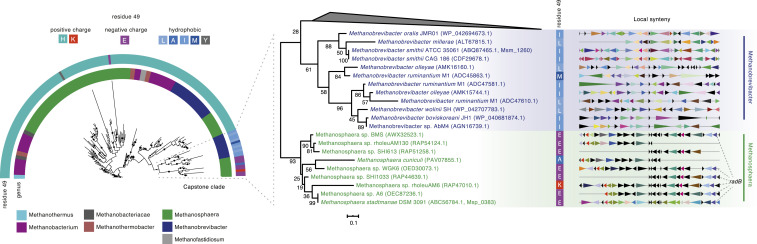
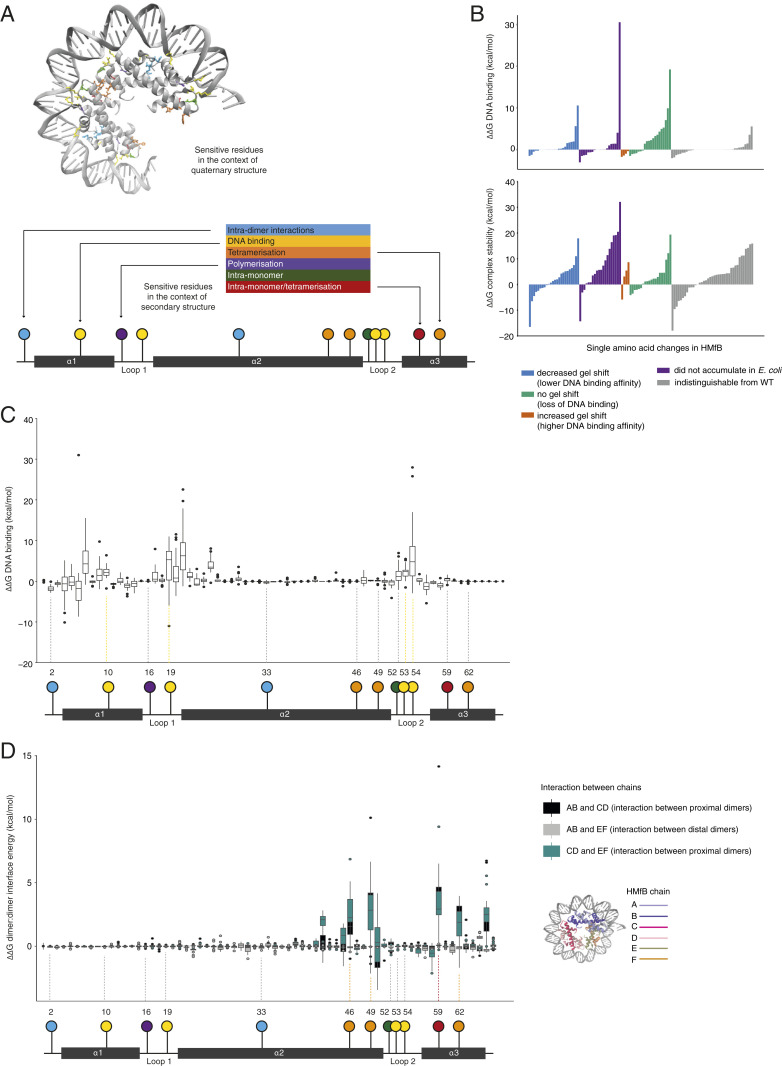
Nucleosomes in eukaryotes act as platforms for the dynamic integration of epigenetic information. Posttranslational modifications are reversibly added or removed and core histones exchanged for paralogous variants, in concert with changing demands on transcription and genome accessibility. Histones are also common in archaea. Their role in genome regulation, however, and the capacity of individual paralogs to assemble into histone-DNA complexes with distinct properties remain poorly understood. Here, we combine structural modeling with phylogenetic analysis to shed light on archaeal histone paralogs, their evolutionary history, and capacity to generate combinatorial chromatin states through hetero-oligomeric assembly. Focusing on the human commensal as a model archaeal system, we show that the heteromeric complexes that can be assembled from its seven histone paralogs vary substantially in DNA binding affinity and tetramer stability. Using molecular dynamics simulations, we go on to identify unique paralogs in and that are characterized by unstable interfaces between dimers. We propose that these paralogs act as capstones that prevent stable tetramer formation and extension into longer oligomers characteristic of model archaeal histones. Importantly, we provide evidence from phylogeny and genome architecture that these capstones, as well as other paralogs in the Methanobacteriales, have been maintained for hundreds of millions of years following ancient duplication events. Taken together, our findings indicate that at least some archaeal histone paralogs have evolved to play distinct and conserved functional roles, reminiscent of eukaryotic histone variants. We conclude that combinatorially complex histone-based chromatin is not restricted to eukaryotes and likely predates their emergence.
真核生物中的核小体作为动态整合表观遗传信息的平台。翻译后修饰可被可逆地添加或去除,核心组蛋白可被同源变体交换,以适应转录和基因组可及性的变化需求。组蛋白也存在于古菌中。然而,它们在基因组调控中的作用,以及单个同源变体组装具有独特性质的组蛋白-DNA 复合物的能力,仍知之甚少。在这里,我们结合结构建模和系统发育分析,阐明了古菌组蛋白同源变体、它们的进化历史,以及通过异源寡聚体组装产生组合染色质状态的能力。我们以人类共生菌 作为模型古菌系统,表明可以从其七个组蛋白同源变体组装的异源寡聚体复合物在 DNA 结合亲和力和四聚体稳定性方面差异很大。使用分子动力学模拟,我们进一步确定了 和 中独特的同源变体,它们的二聚体之间的界面不稳定。我们提出,这些同源变体充当顶石,防止稳定的四聚体形成和延伸到具有模型古菌组蛋白特征的更长寡聚体。重要的是,我们从系统发育和基因组结构中提供证据表明,这些顶石以及 Methanobacteriales 中的其他同源变体,在古老的复制事件之后已经被保留了数亿年。总之,我们的发现表明,至少一些古菌组蛋白同源变体已经进化到发挥独特且保守的功能作用,类似于真核组蛋白变体。我们得出结论,组合复杂的基于组蛋白的染色质不仅限于真核生物,并且可能早于它们的出现而存在。