Department of Molecular Machines and Signaling, Max Planck Institute of Biochemistry, Martinsried, Germany.
Carle Illinois College of Medicine, Champaign, IL, USA.
Nat Chem Biol. 2021 Mar;17(3):272-279. doi: 10.1038/s41589-020-00696-0. Epub 2020 Dec 7.
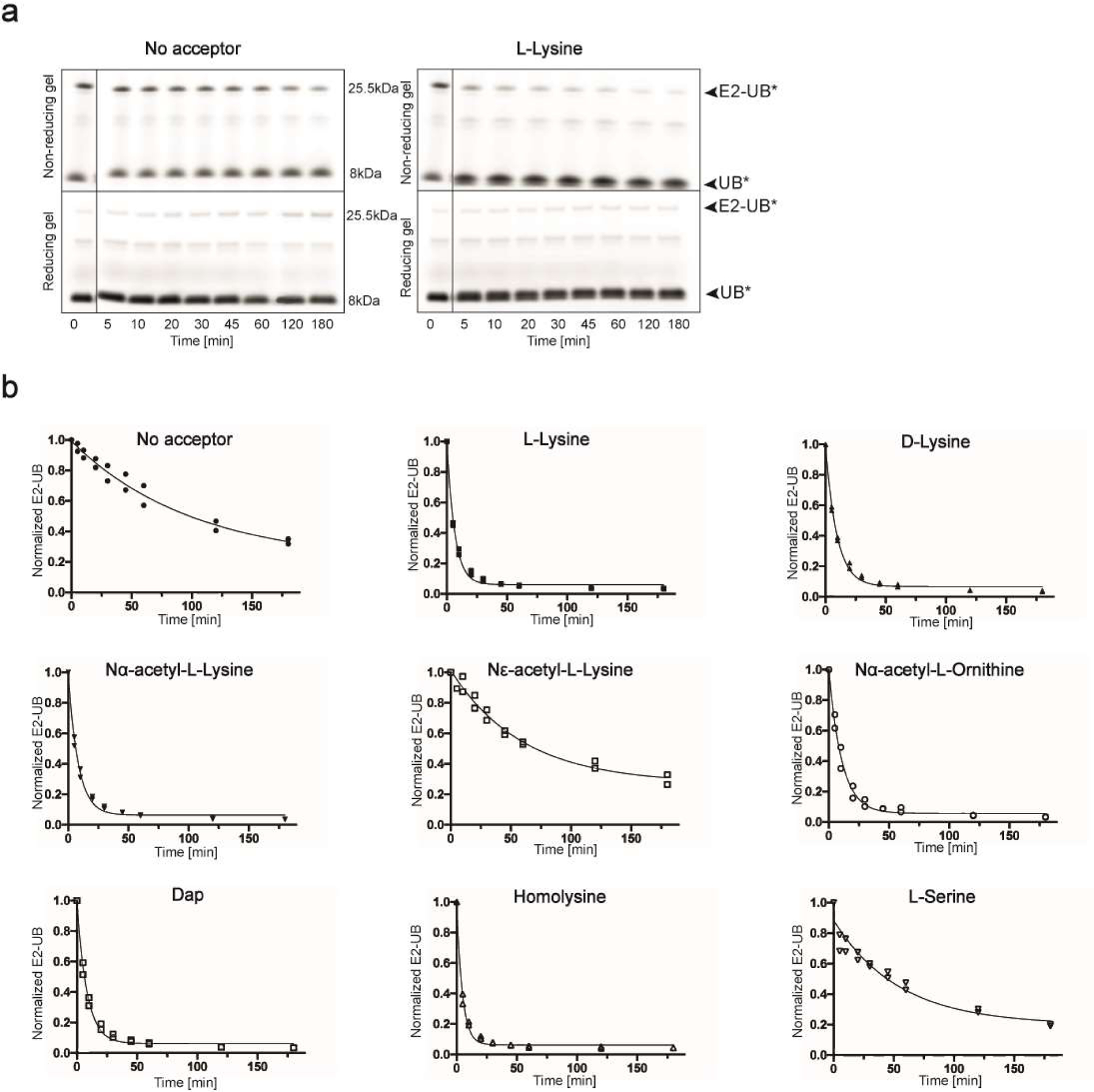
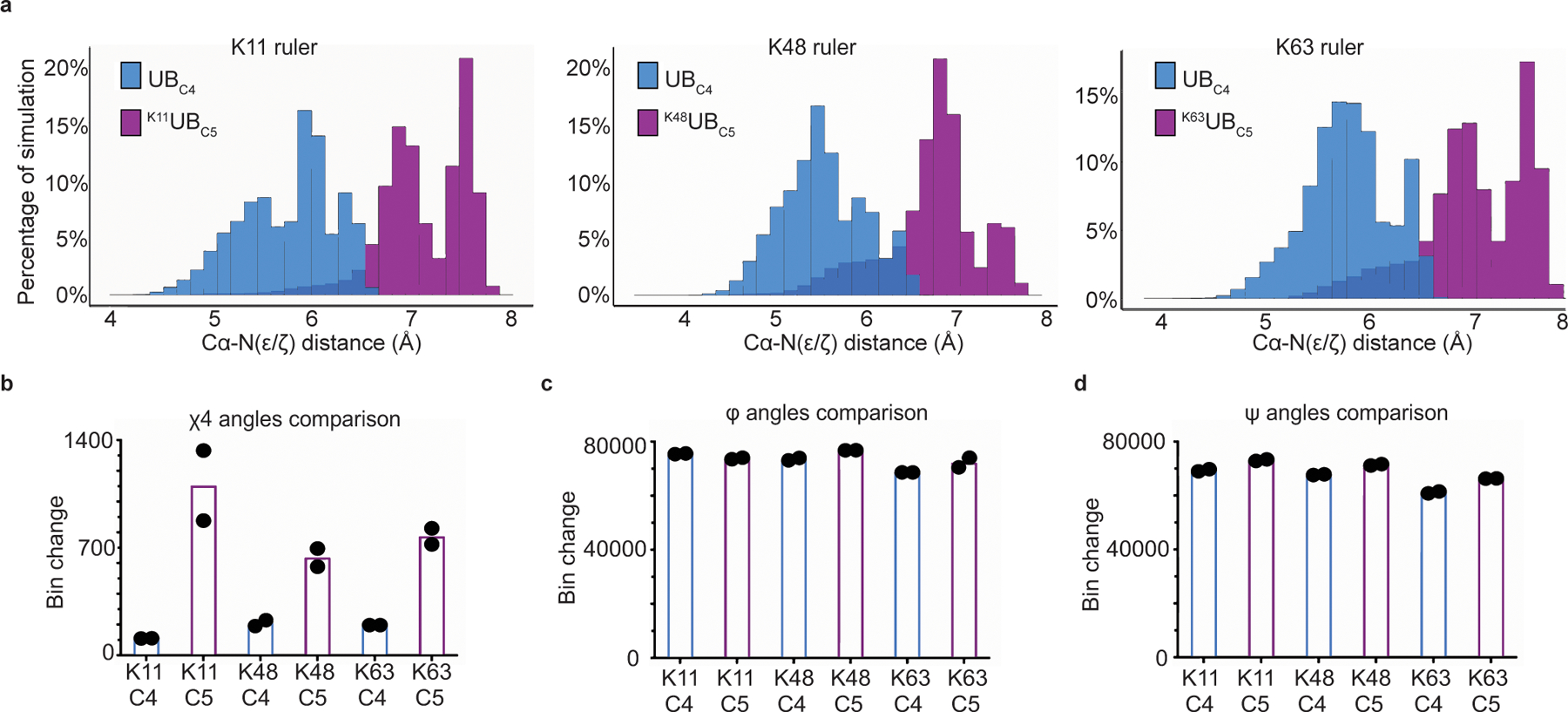
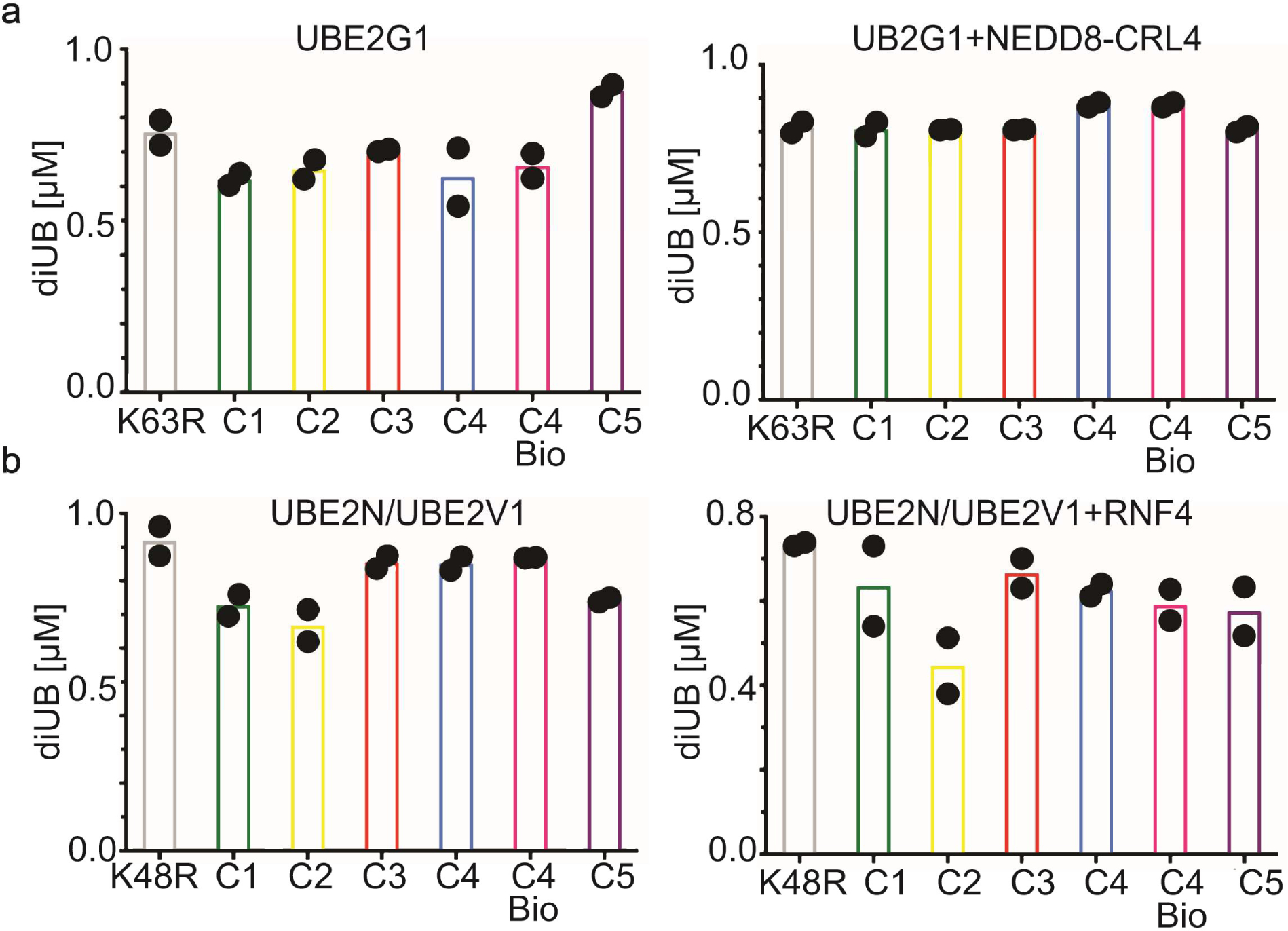
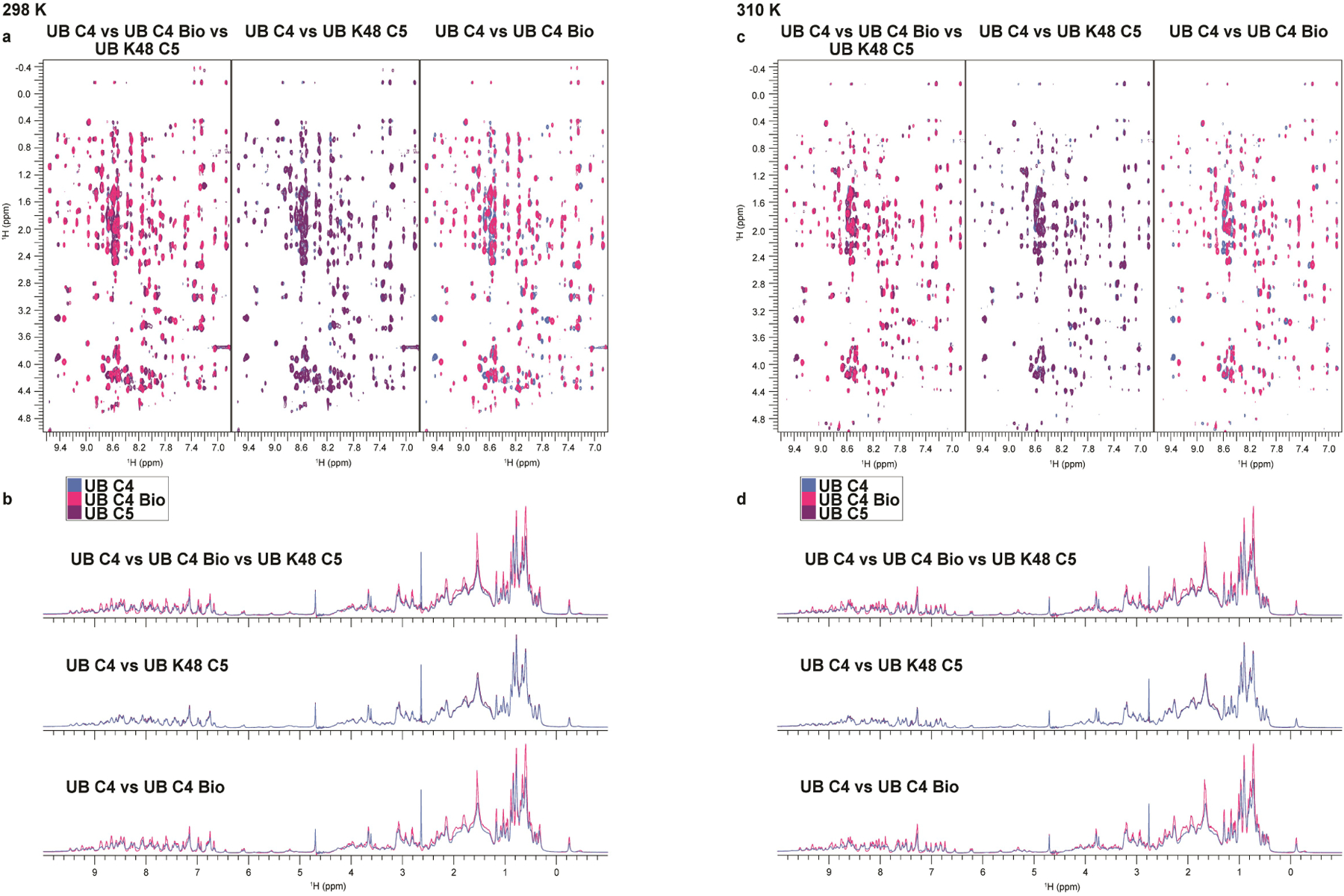
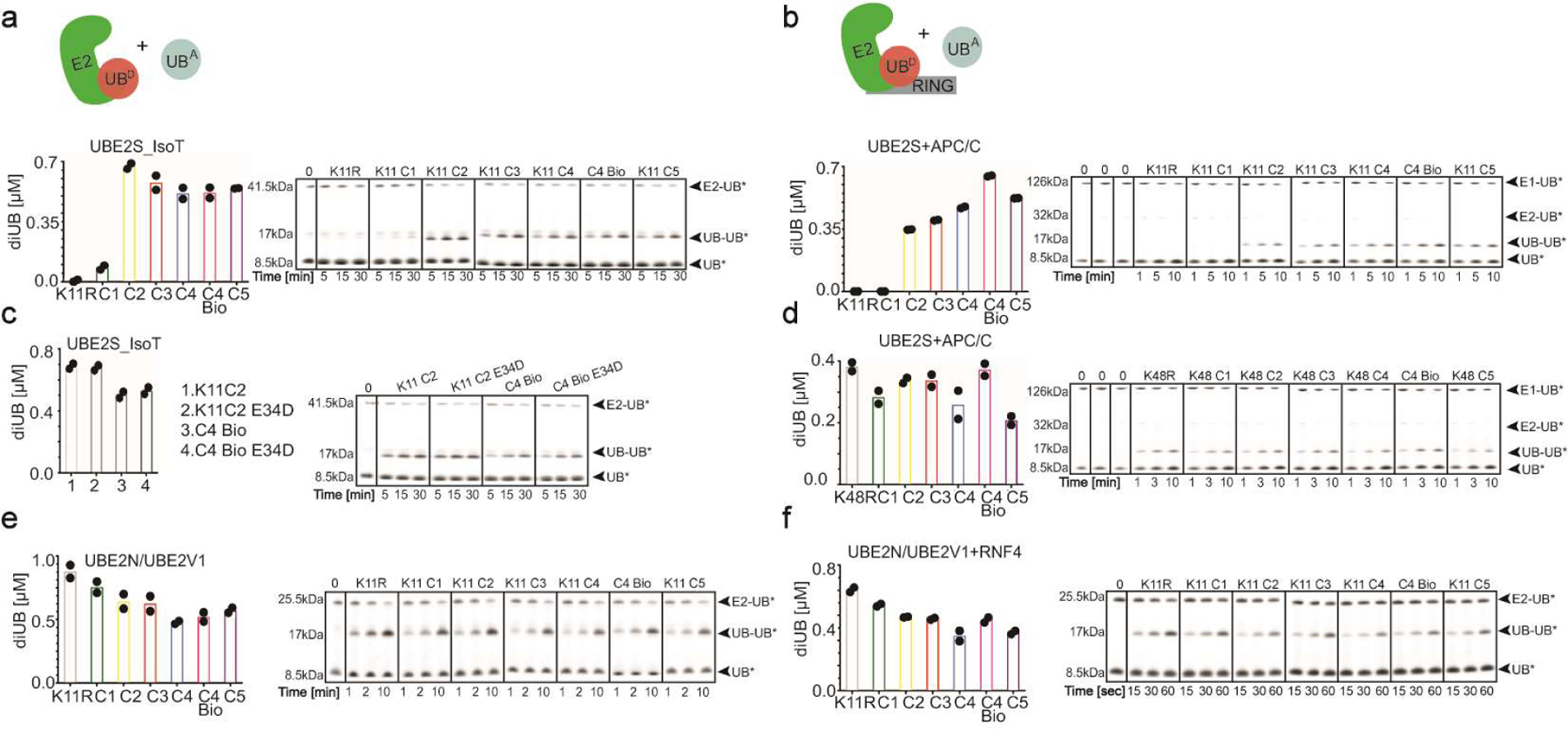
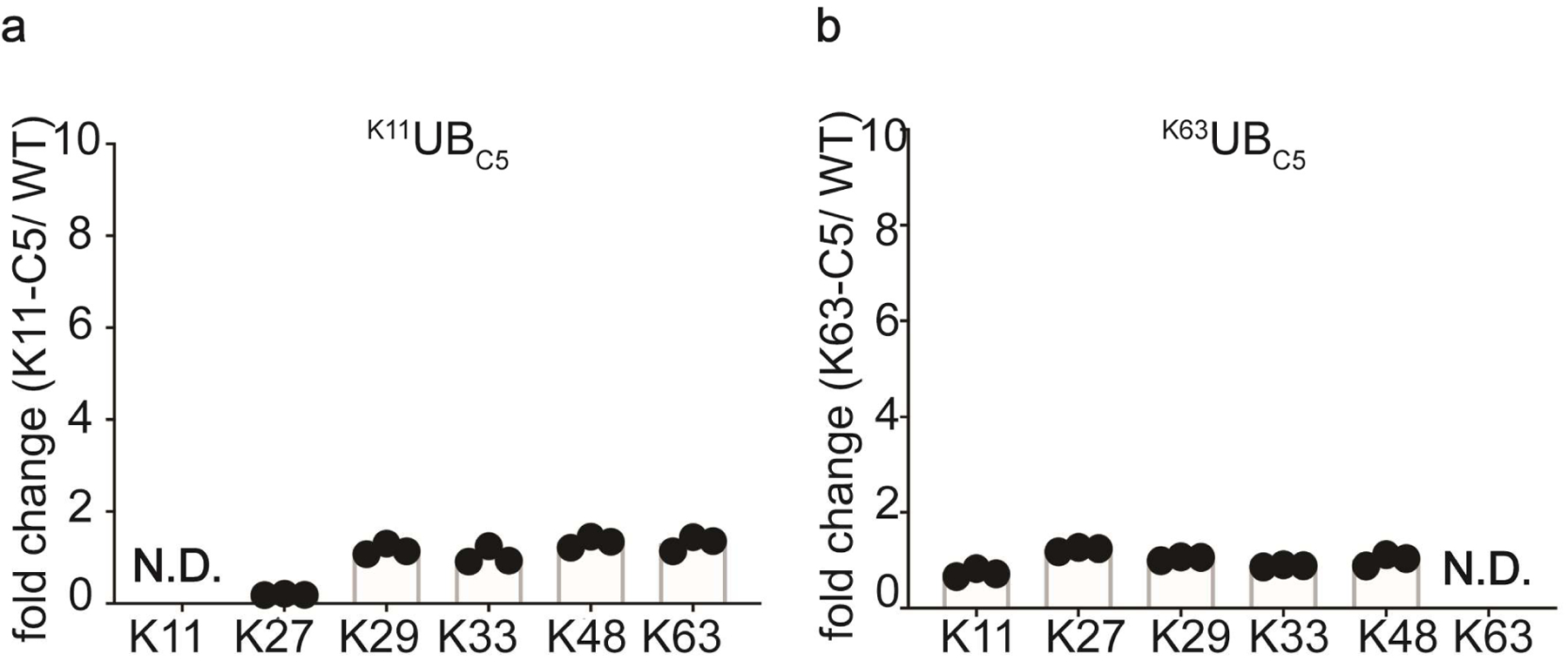
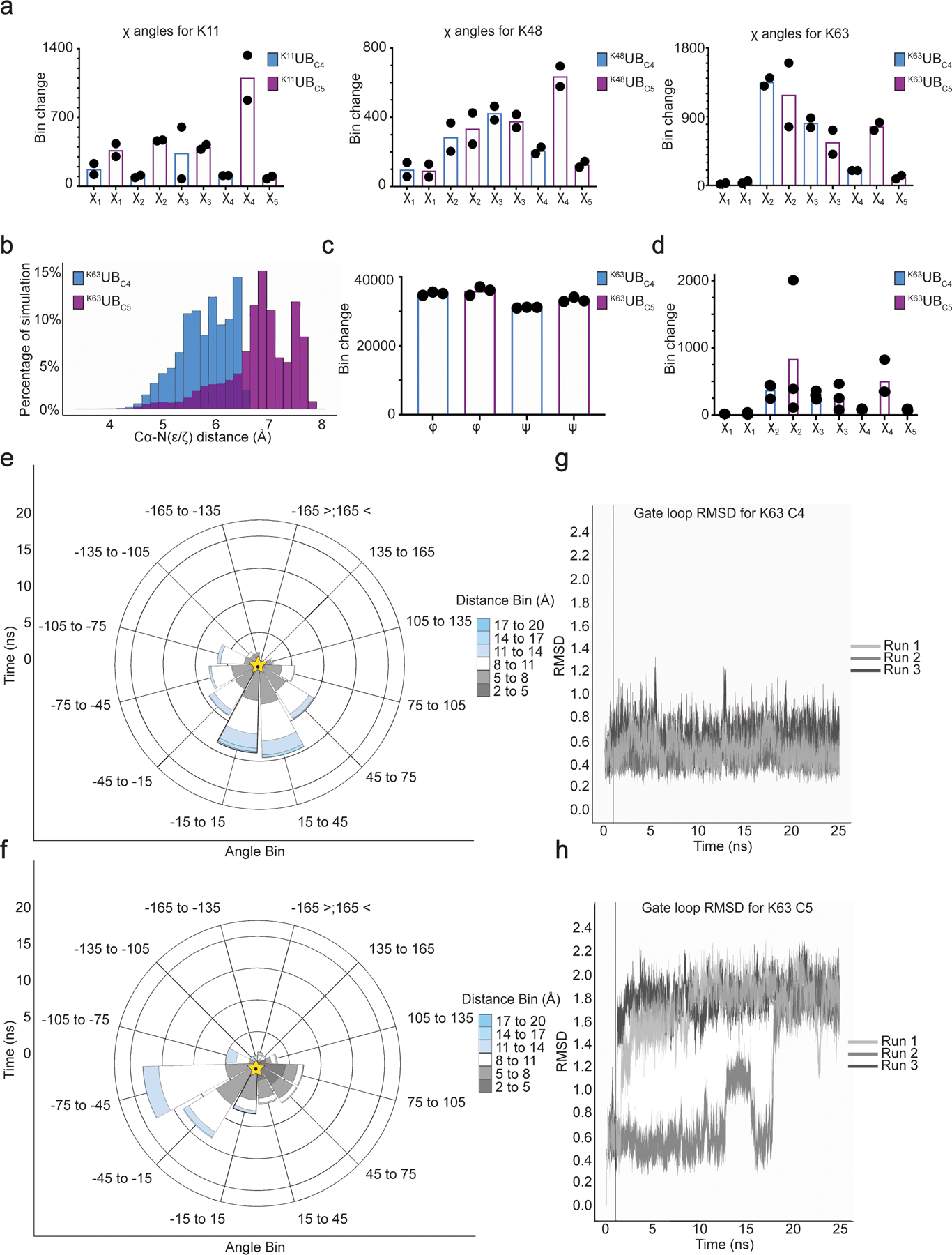
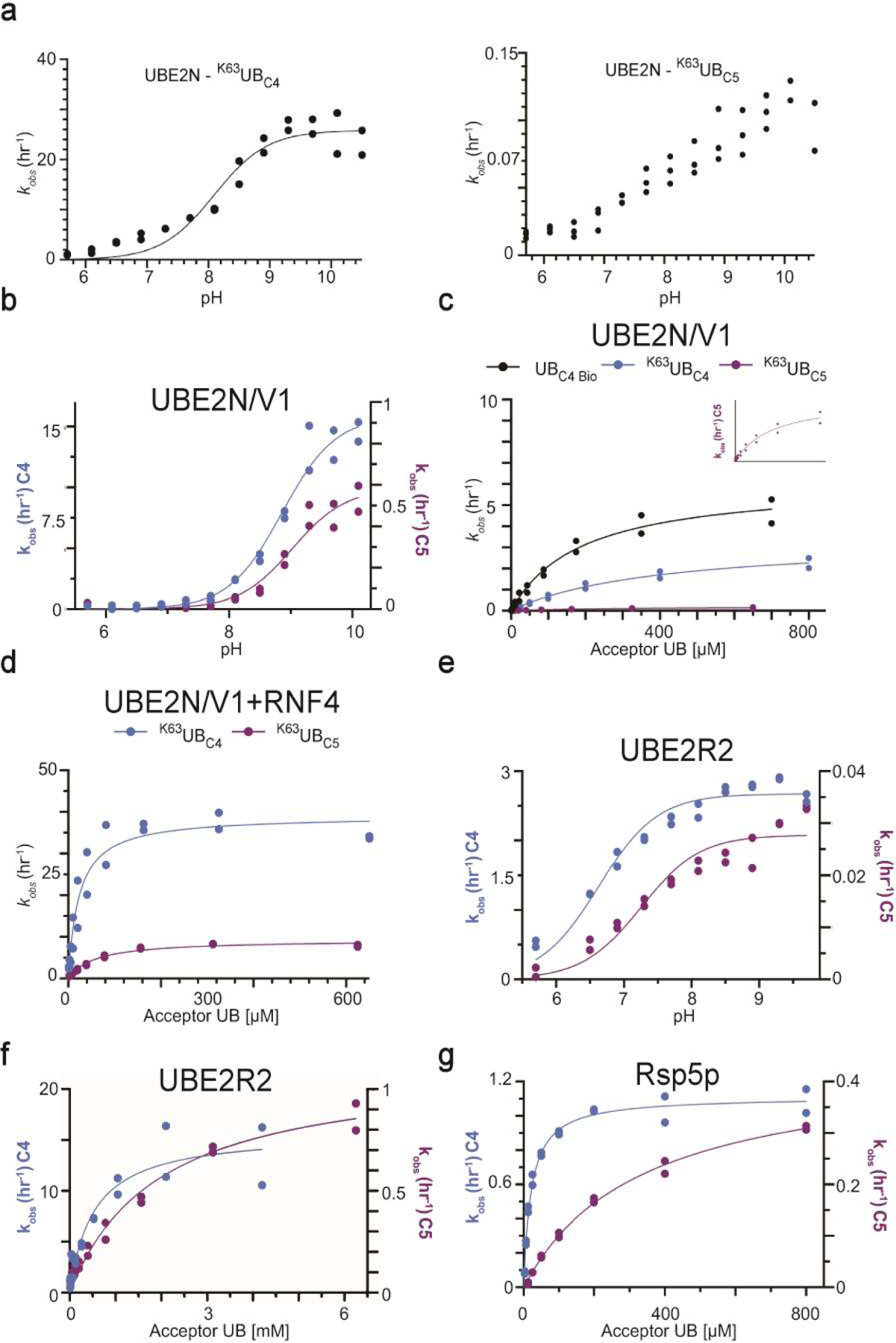
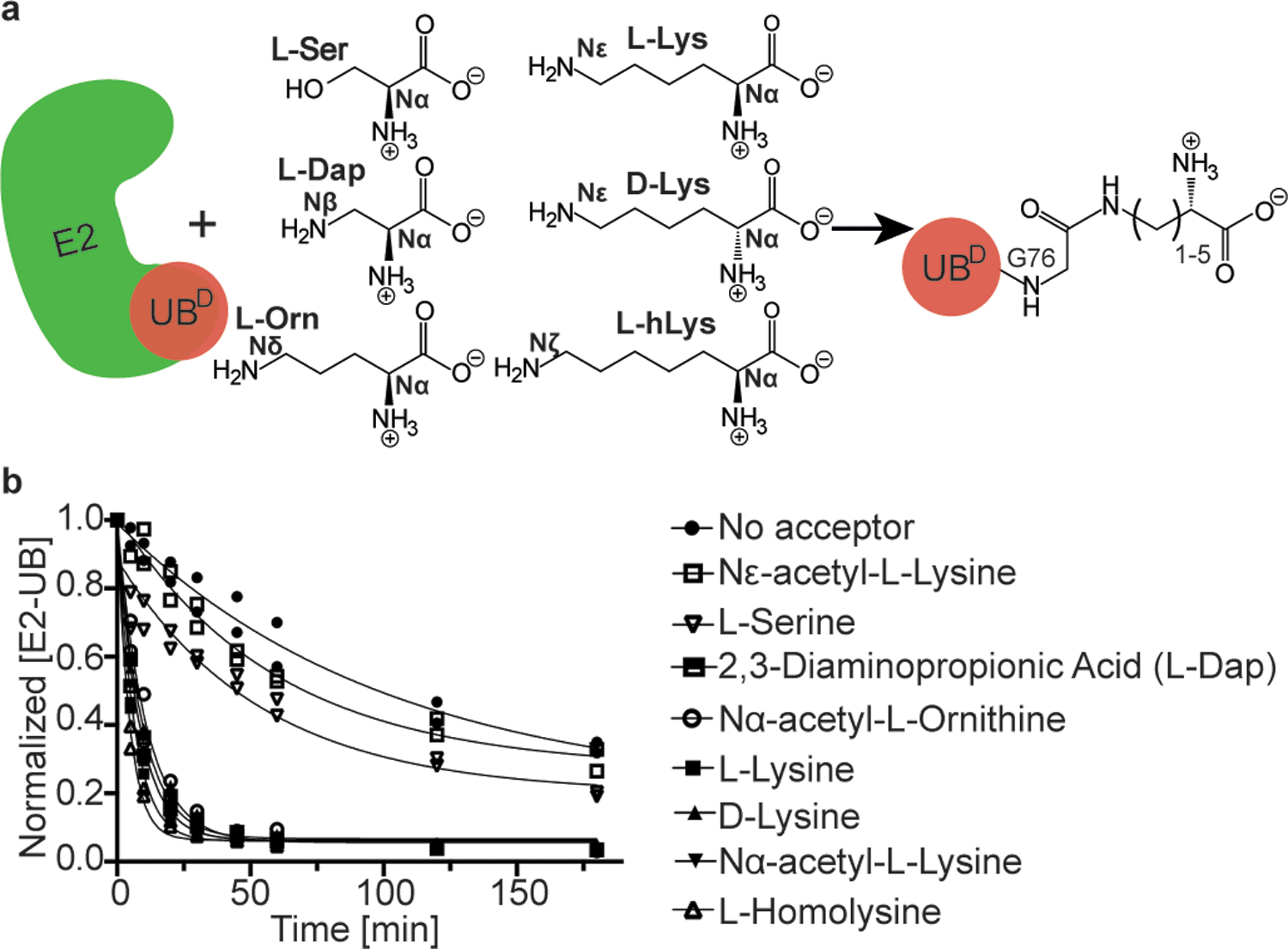
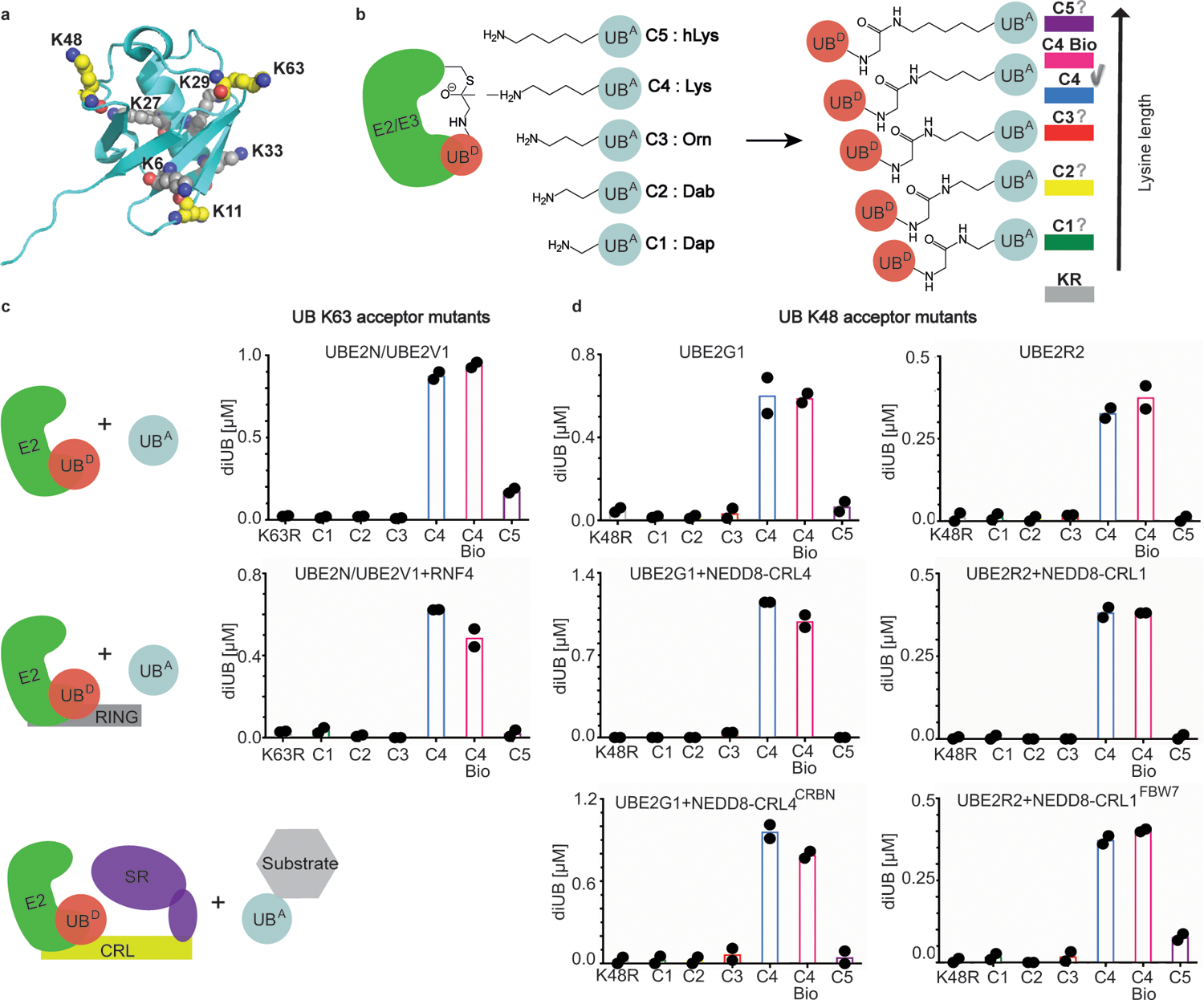
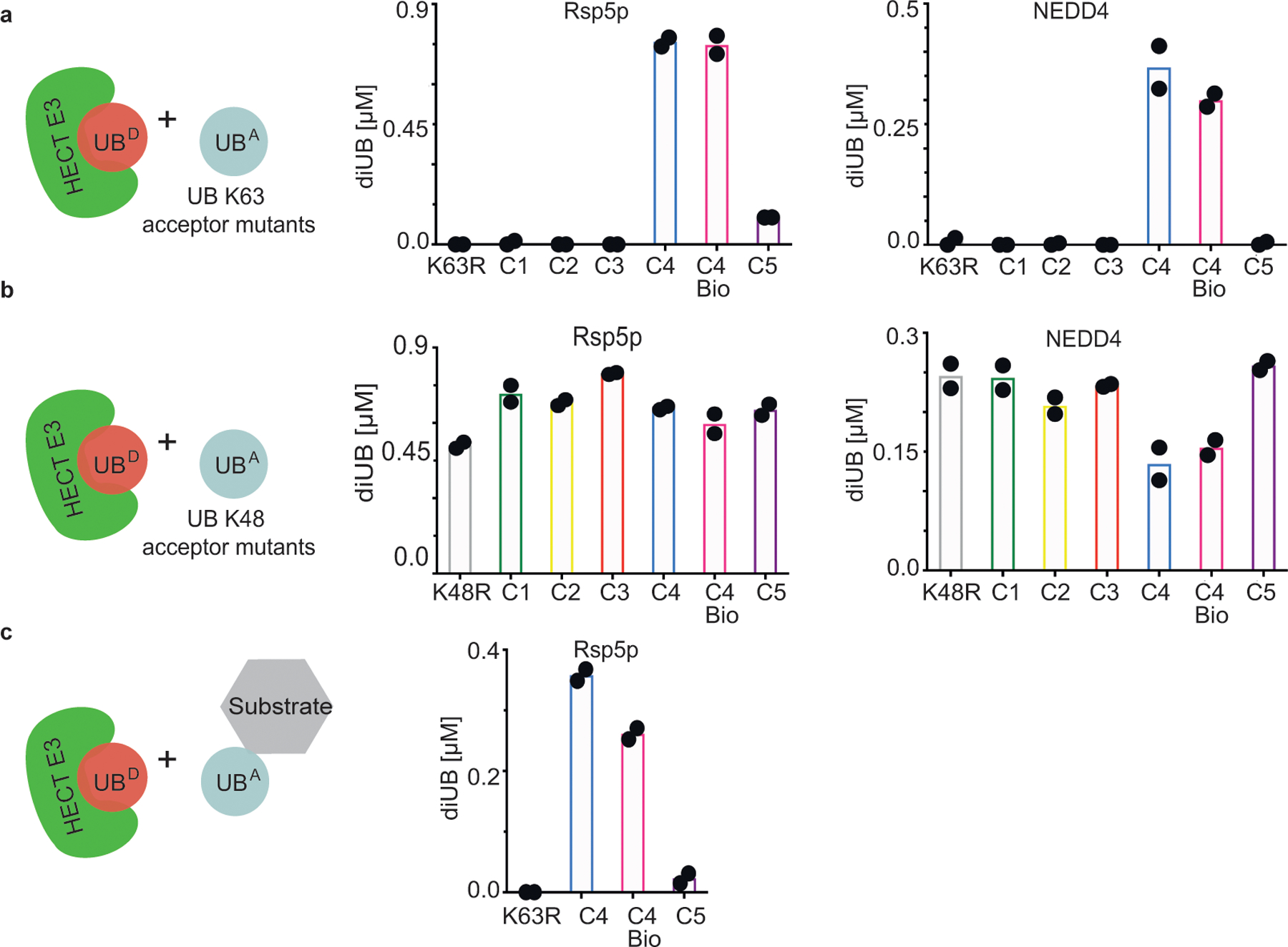
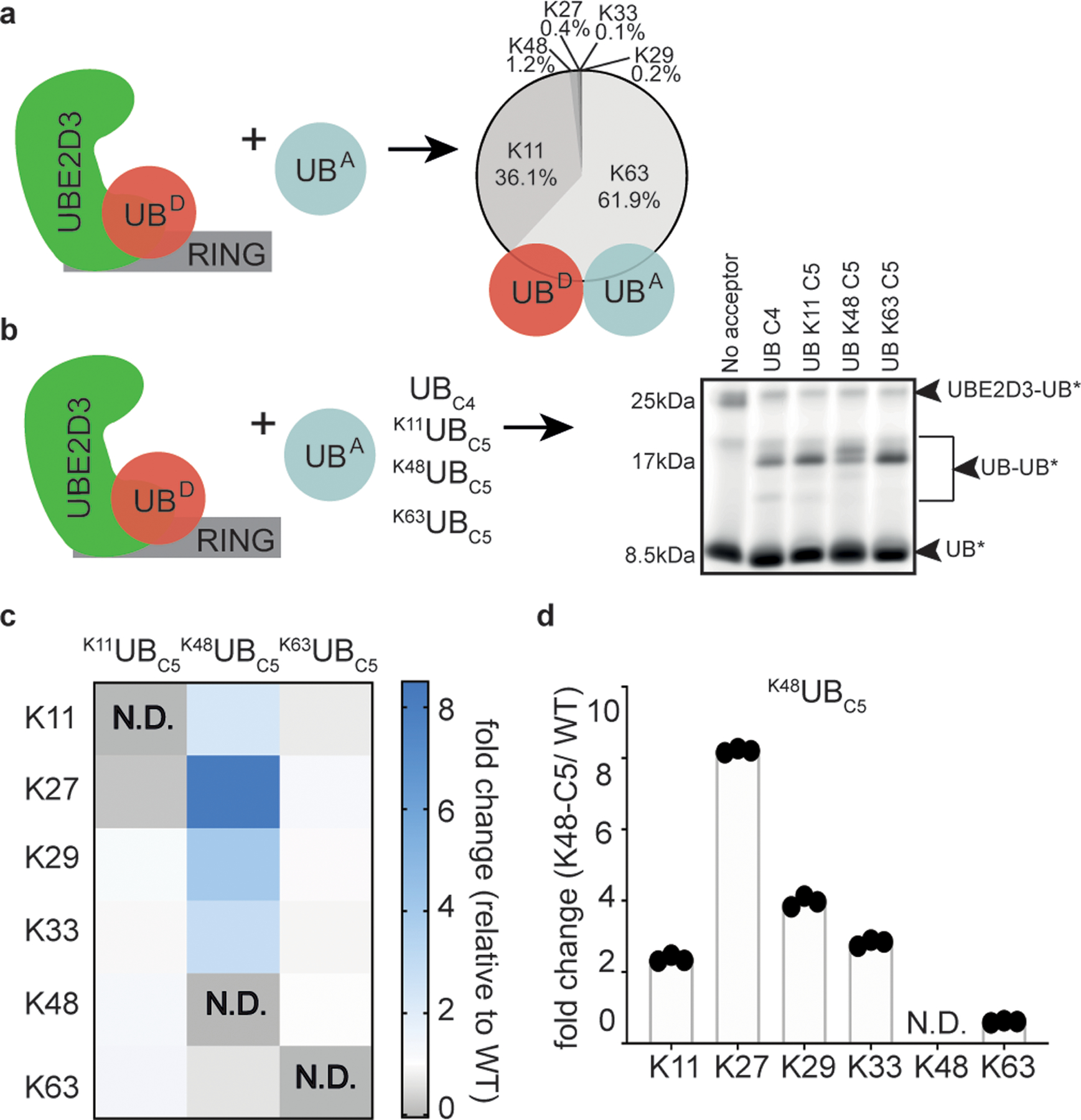
Virtually all aspects of cell biology are regulated by a ubiquitin code where distinct ubiquitin chain architectures guide the binding events and itineraries of modified substrates. Various combinations of E2 and E3 enzymes accomplish chain formation by forging isopeptide bonds between the C terminus of their transiently linked donor ubiquitin and a specific nucleophilic amino acid on the acceptor ubiquitin, yet it is unknown whether the fundamental feature of most acceptors-the lysine side chain-affects catalysis. Here, use of synthetic ubiquitins with non-natural acceptor site replacements reveals that the aliphatic side chain specifying reactive amine geometry is a determinant of the ubiquitin code, through unanticipated and complex reliance of many distinct ubiquitin-carrying enzymes on a canonical acceptor lysine.
细胞生物学的几乎所有方面都受到泛素编码的调控,其中不同的泛素链结构指导修饰底物的结合事件和轨迹。各种 E2 和 E3 酶通过在它们短暂连接的供体泛素的 C 末端和受体泛素上的特定亲核氨基酸之间形成异肽键来完成链的形成,然而,目前尚不清楚大多数受体-赖氨酸侧链-是否影响催化的基本特征。在这里,使用具有非天然受体位点取代的合成泛素揭示了指定反应性胺几何形状的脂肪侧链是泛素编码的决定因素,这是通过许多不同的携带泛素的酶对典型受体赖氨酸的出乎意料和复杂的依赖。