Department of Biochemistry and Biophysics, School of Basic Medical Sciences, Peking University, Beijing, China.
Department of Structural Biology, Van Andel Institute, Grand Rapids, United States.
Elife. 2020 Dec 15;9:e62163. doi: 10.7554/eLife.62163.
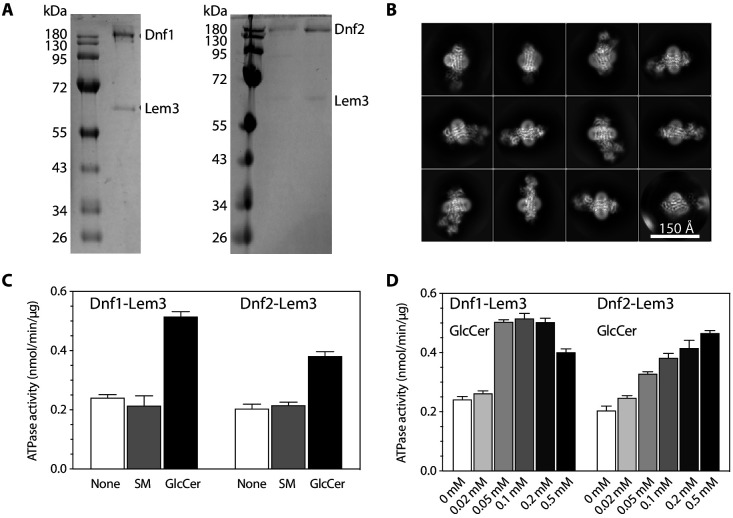
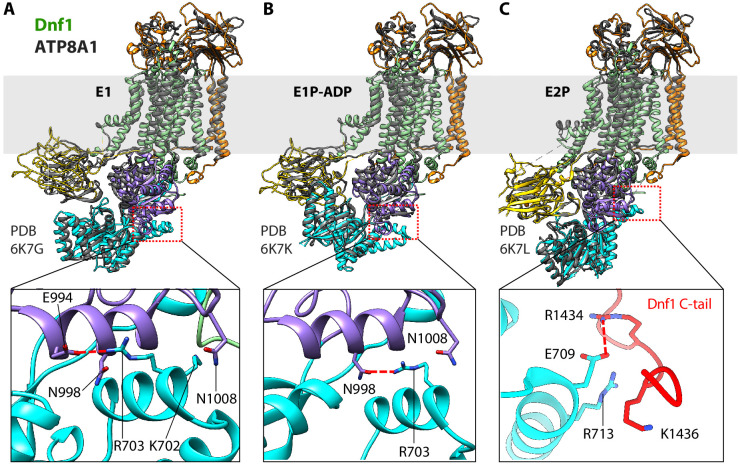
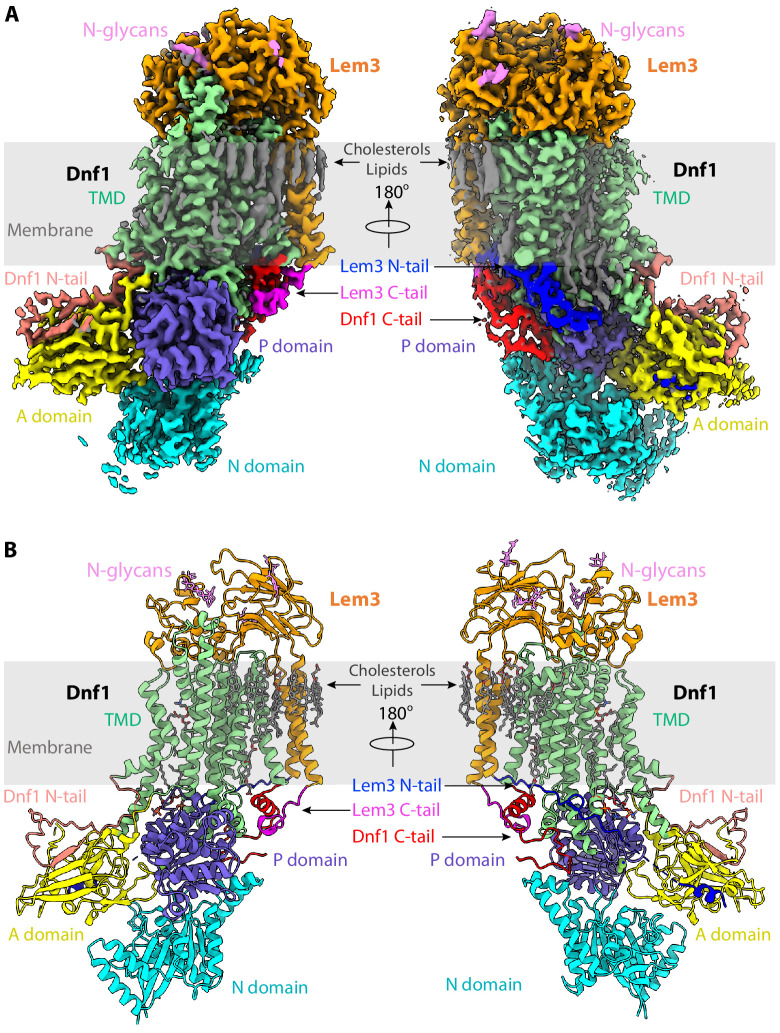
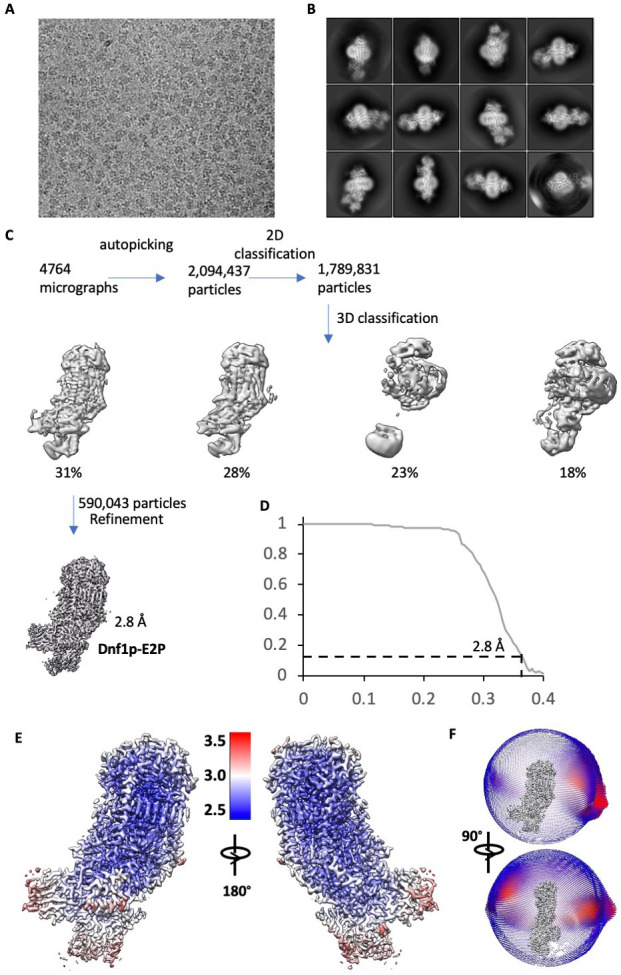
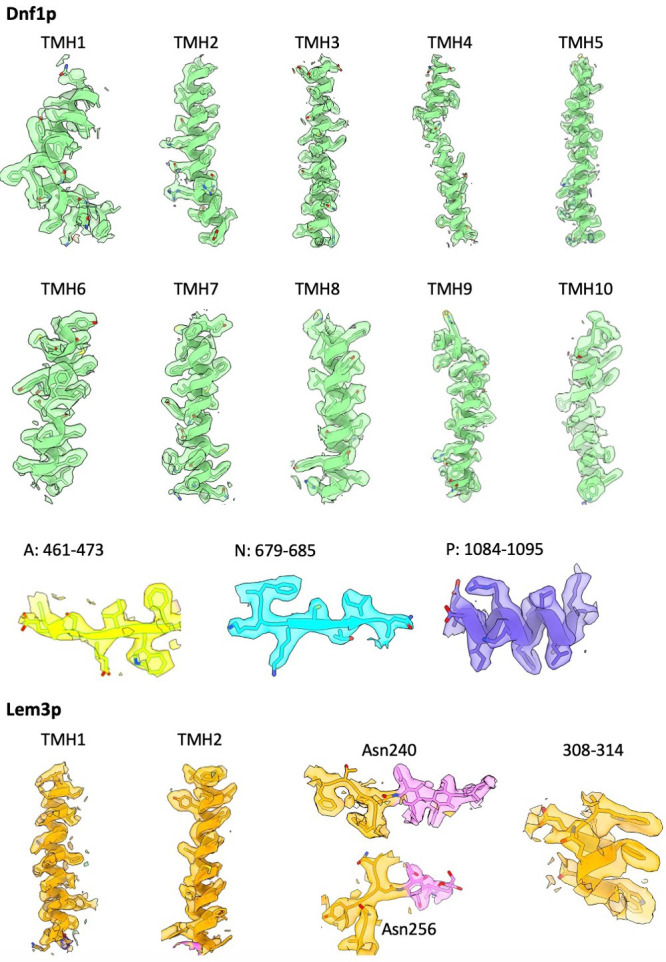
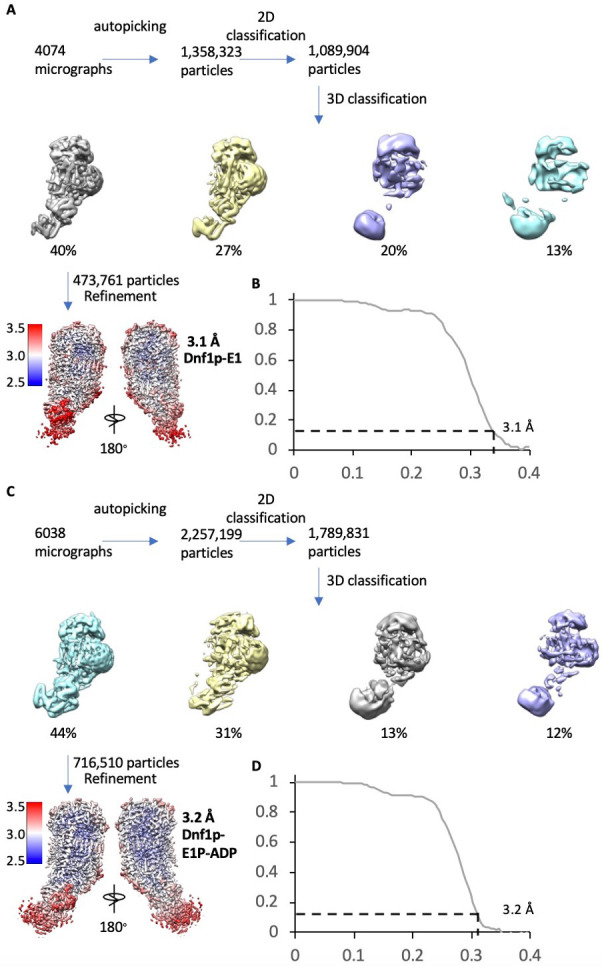
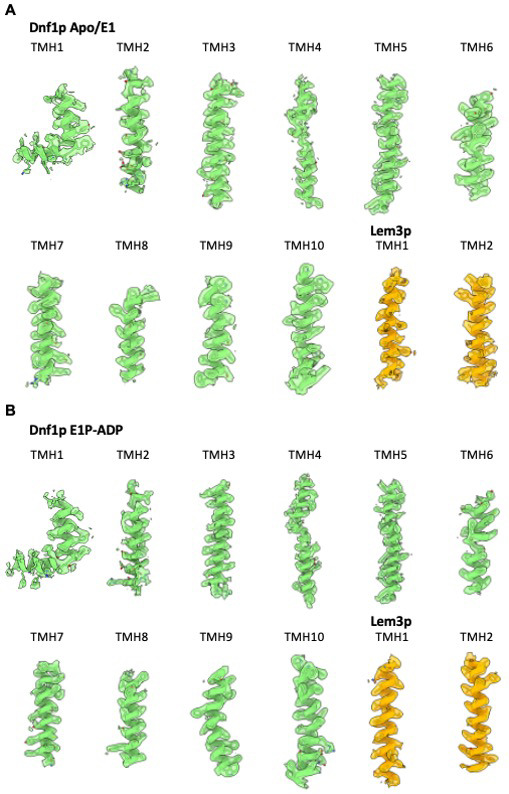
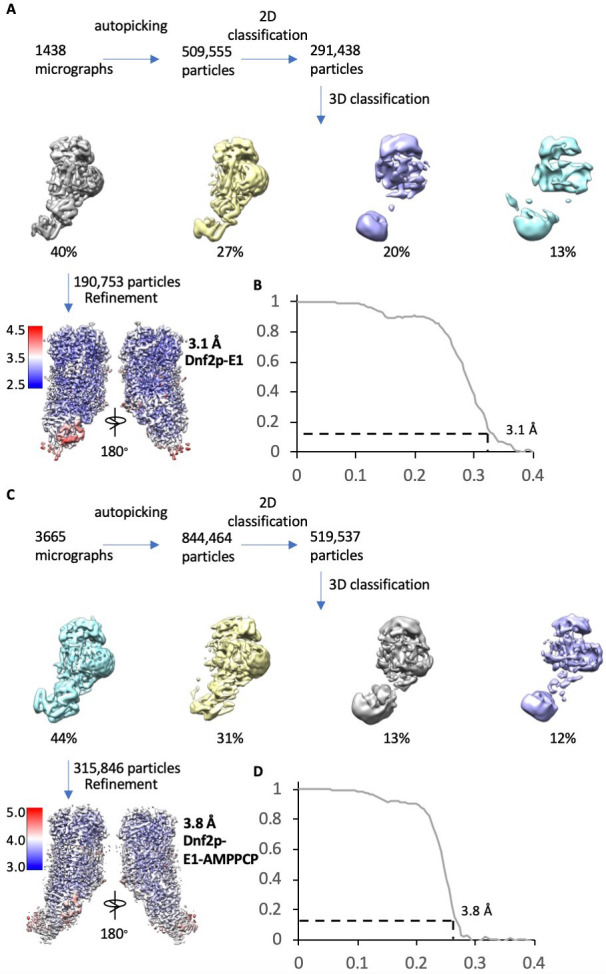
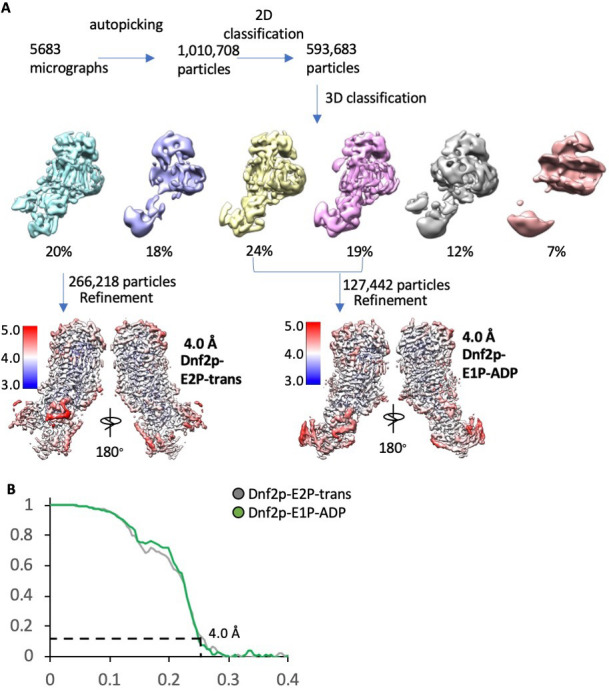
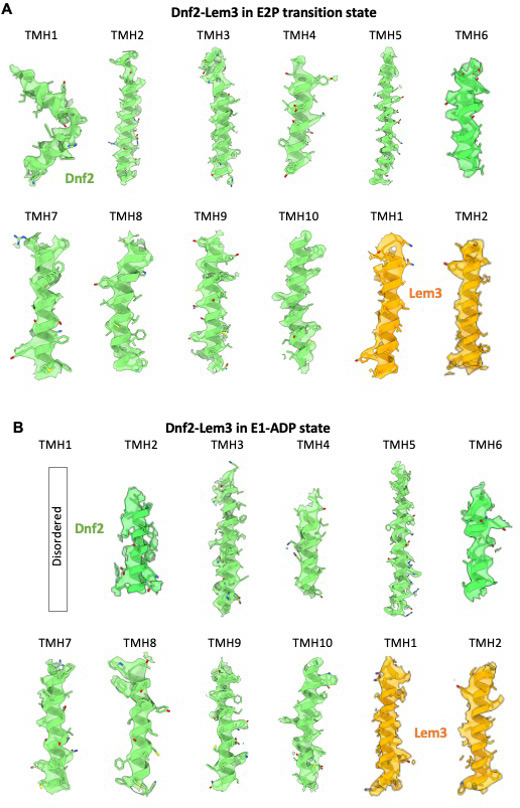
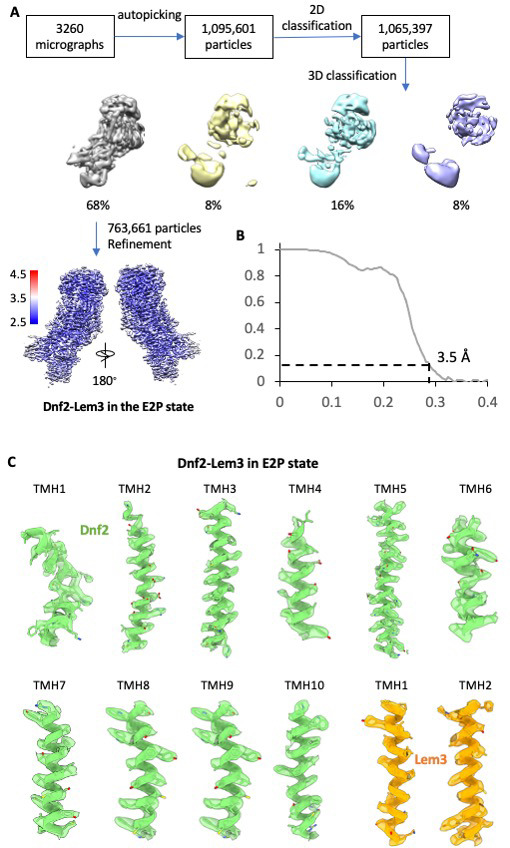
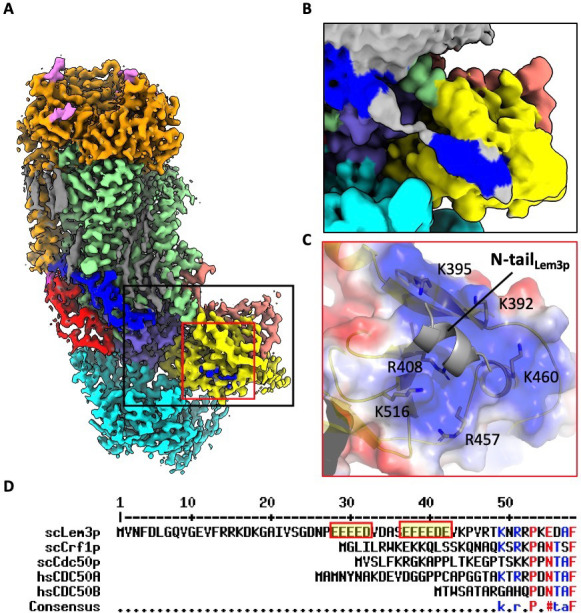
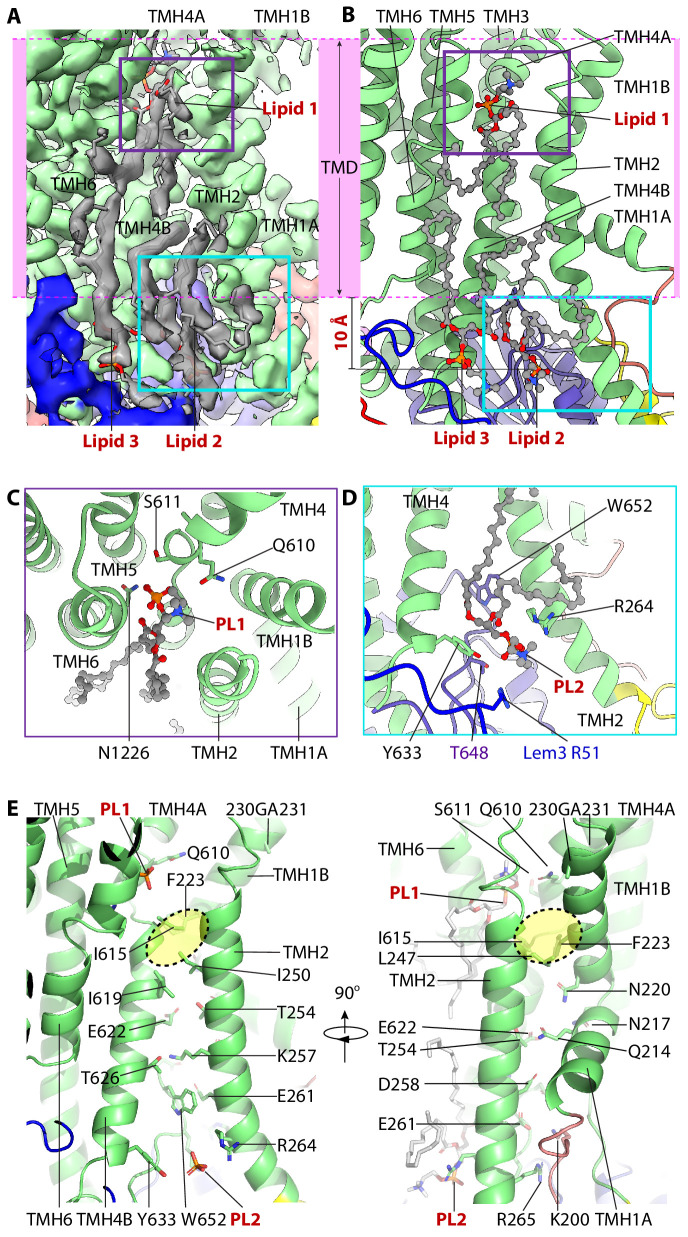
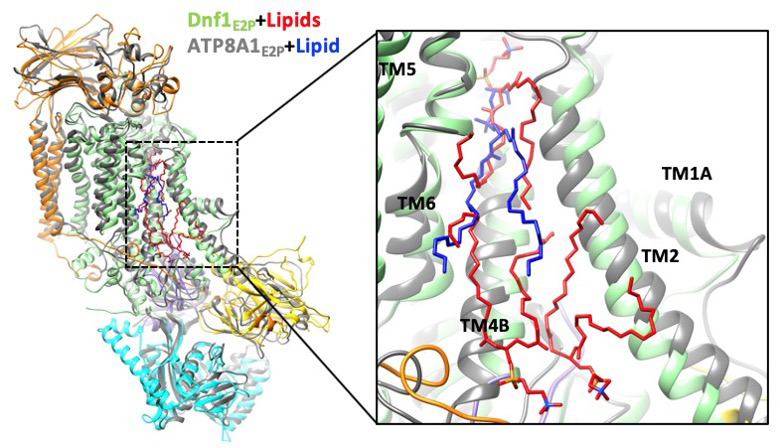
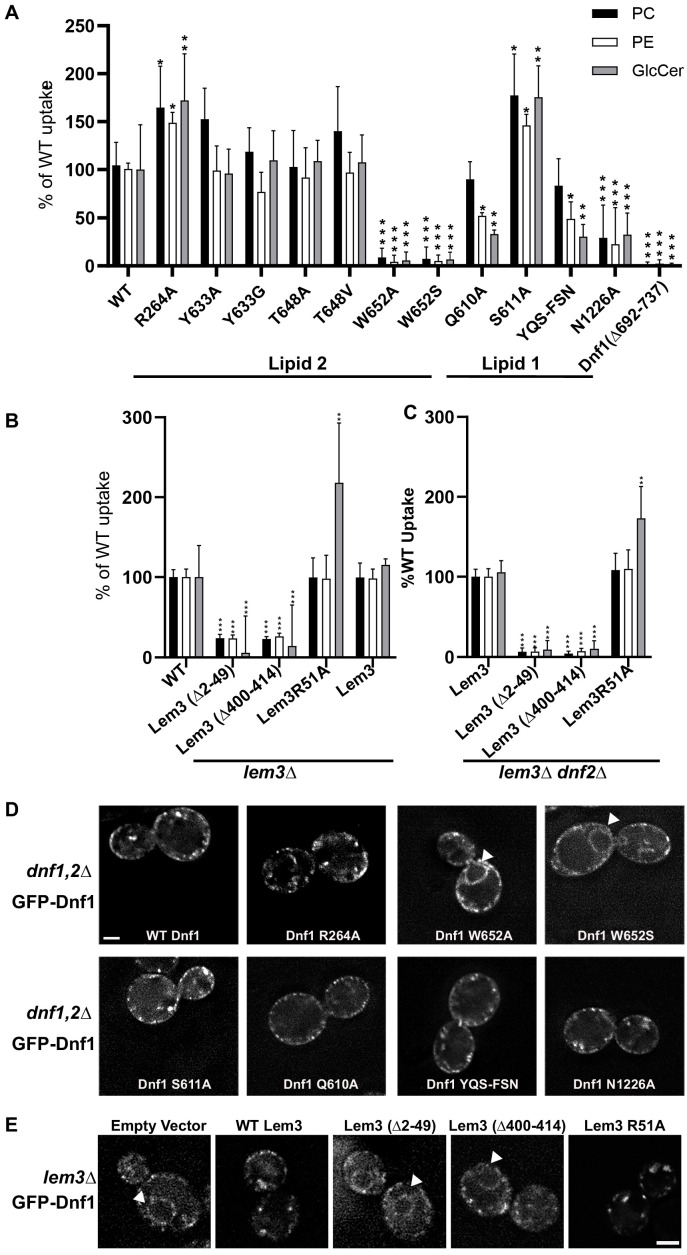
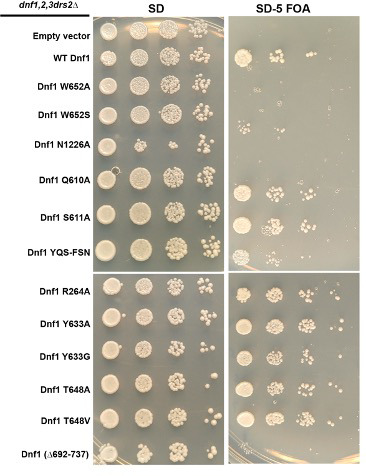
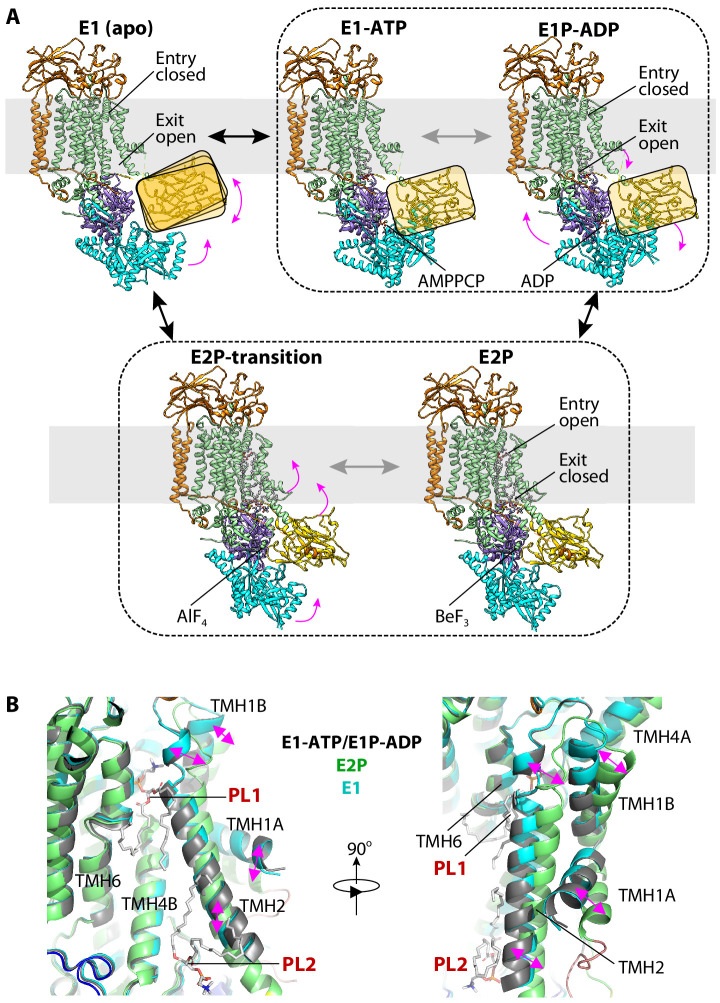
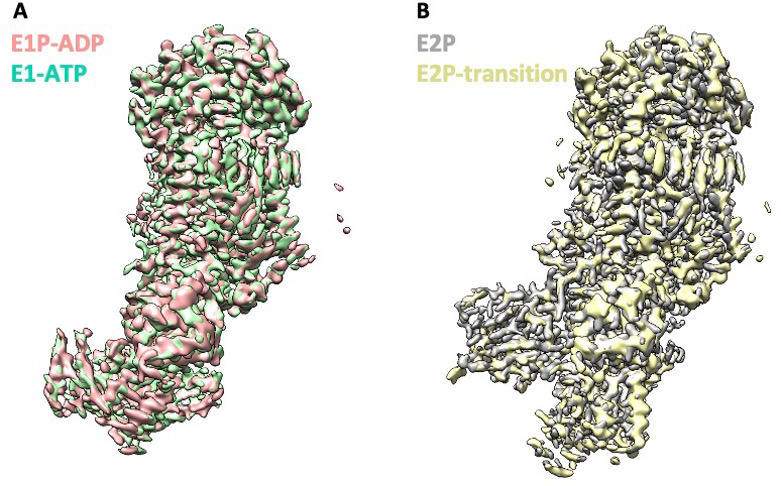
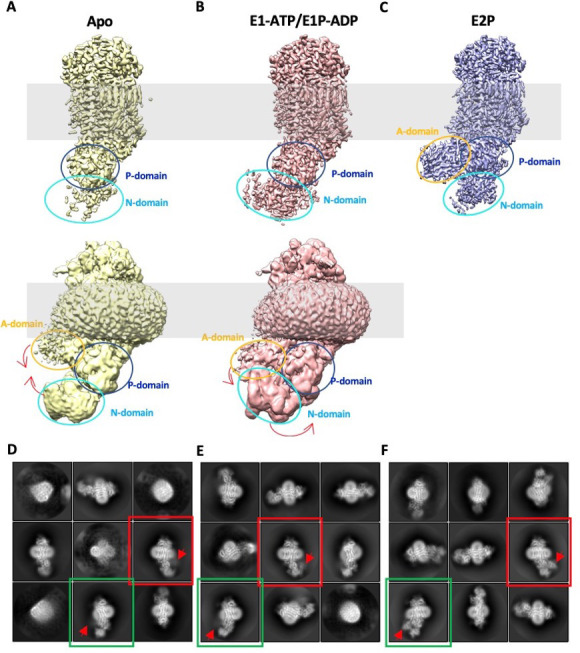
The P4 ATPases use ATP hydrolysis to transport large lipid substrates across lipid bilayers. The structures of the endosome- and Golgi-localized phosphatidylserine flippases-such as the yeast Drs2 and human ATP8A1-have recently been reported. However, a substrate-binding site on the cytosolic side has not been found, and the transport mechanisms of P4 ATPases with other substrates are unknown. Here, we report structures of the Dnf1-Lem3 and Dnf2-Lem3 complexes. We captured substrate phosphatidylcholine molecules on both the exoplasmic and cytosolic sides and found that they have similar structures. Unexpectedly, Lem3 contributes to substrate binding. The conformational transitions of these phosphatidylcholine transporters match those of the phosphatidylserine transporters, suggesting a conserved mechanism among P4 ATPases. Dnf1/Dnf2 have a unique P domain helix-turn-helix insertion that is important for function. Therefore, P4 ATPases may have retained an overall transport mechanism while evolving distinct features for different lipid substrates.
P4 ATP 酶利用 ATP 水解将大的脂质底物跨膜转运。最近报道了内体和高尔基体定位的磷脂酰丝氨酸翻转酶(如酵母 Drs2 和人 ATP8A1)的结构。然而,尚未发现细胞质侧的底物结合位点,并且其他底物的 P4 ATP 酶的转运机制尚不清楚。在这里,我们报告了 Dnf1-Lem3 和 Dnf2-Lem3 复合物的结构。我们在质外侧和细胞质侧捕获了底物磷脂酰胆碱分子,并发现它们具有相似的结构。出乎意料的是,Lem3 有助于底物结合。这些磷脂转运蛋白的构象转变与磷脂酰丝氨酸转运蛋白的构象转变相匹配,表明 P4 ATP 酶之间存在保守的机制。Dnf1/Dnf2 具有独特的 P 结构域螺旋-转角-螺旋插入,这对于功能很重要。因此,P4 ATP 酶可能在保留不同脂质底物的独特特征的同时保留了总体转运机制。