Division of Biological Sciences, University of California, San Diego, La Jolla, United States.
Elife. 2021 Feb 2;10:e60200. doi: 10.7554/eLife.60200.
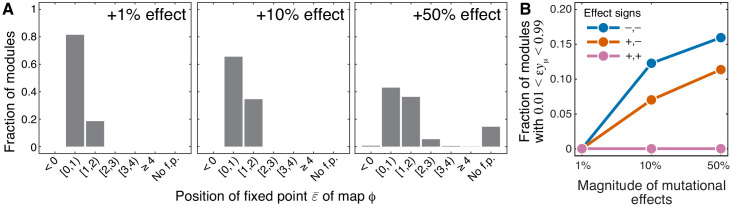
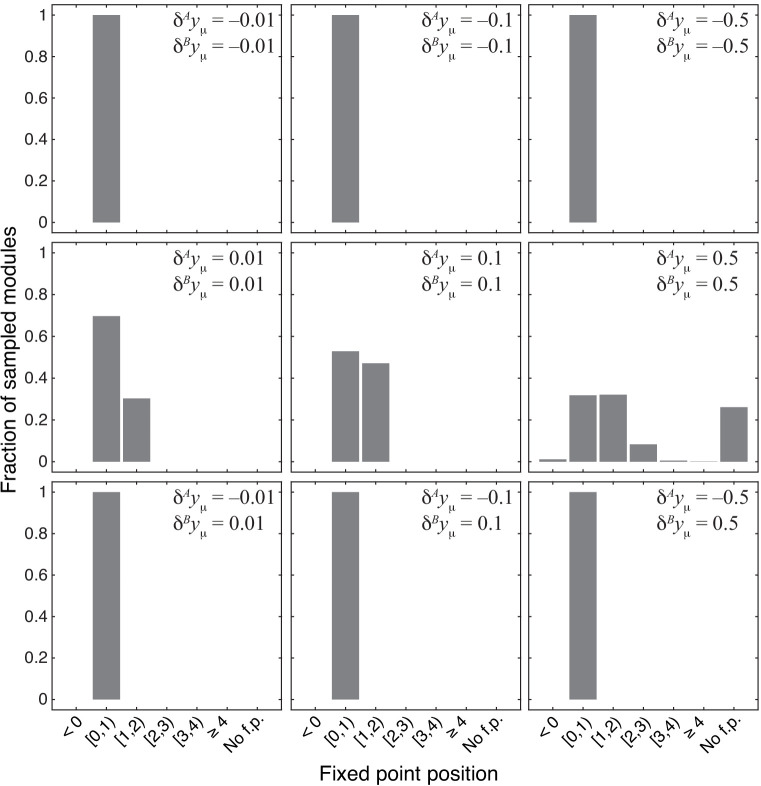
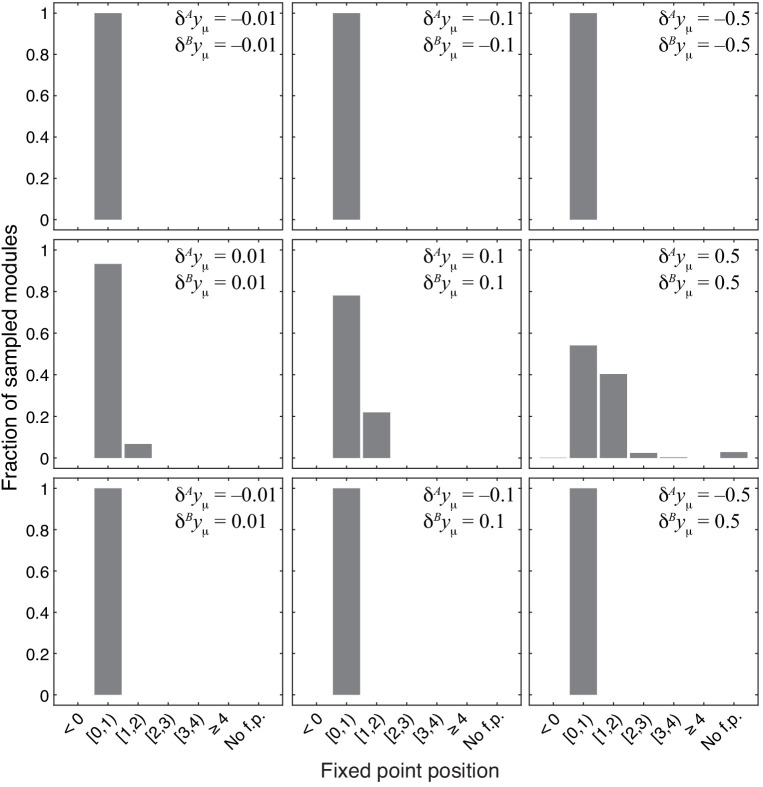
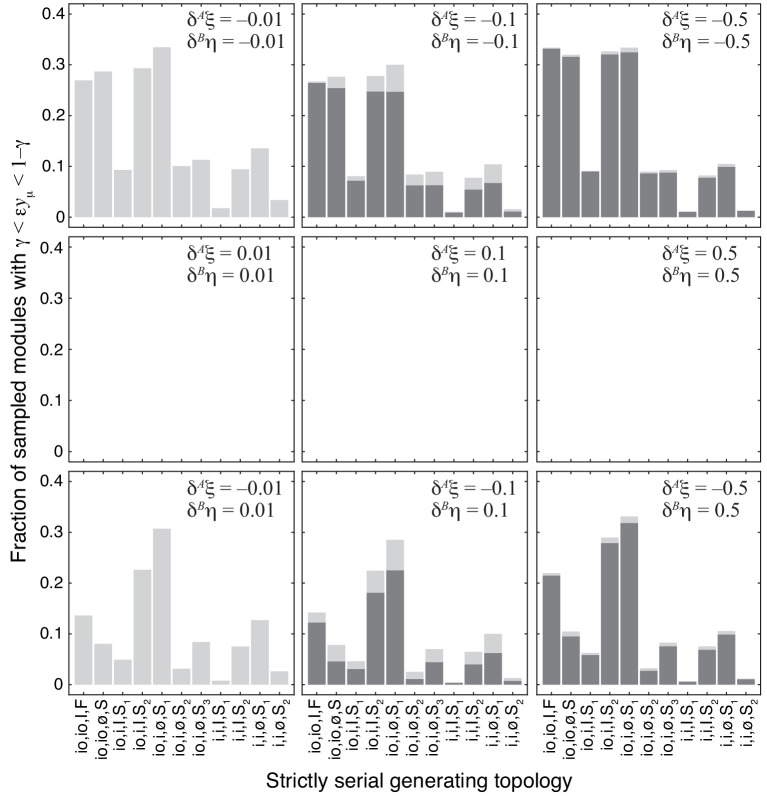
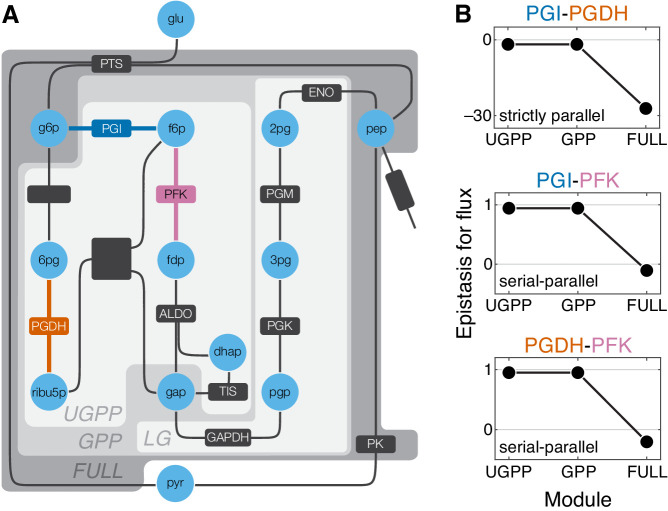
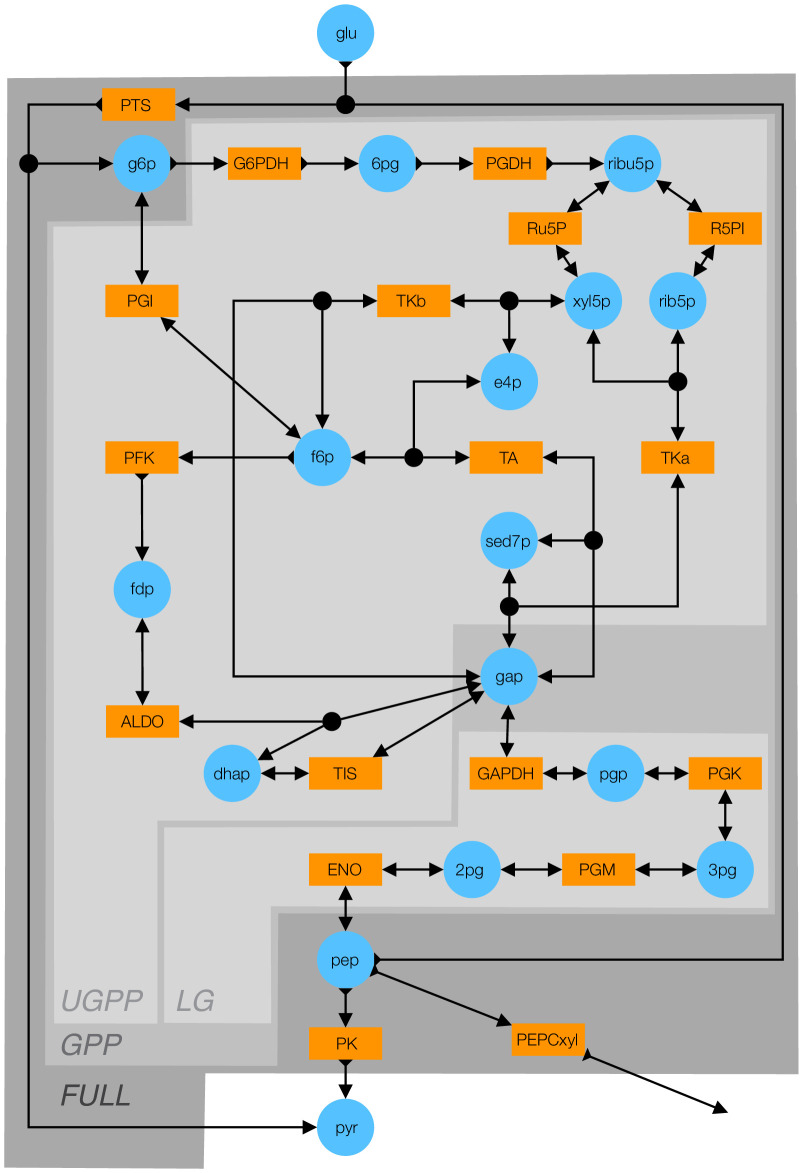
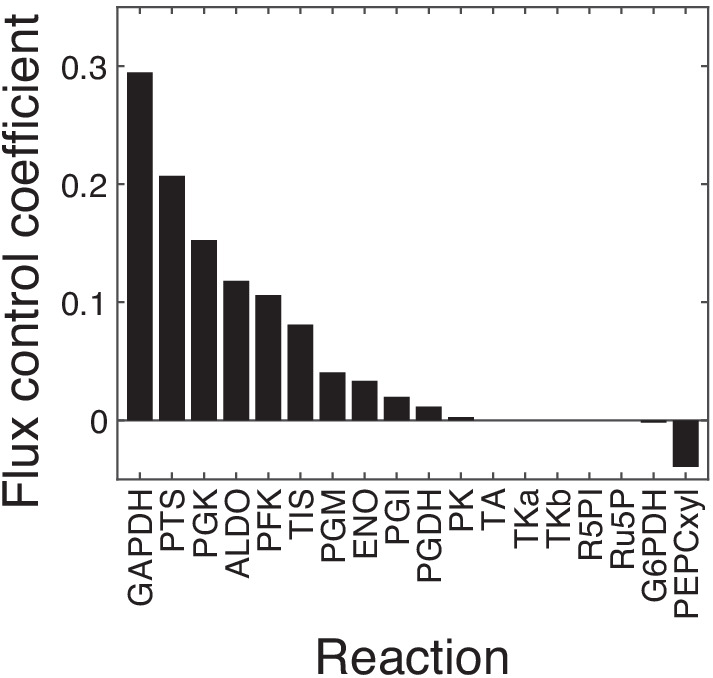
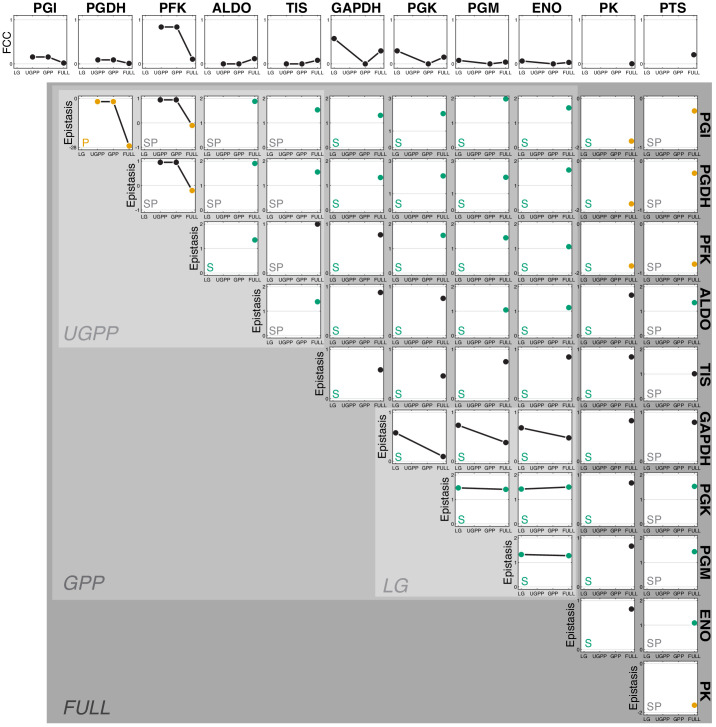
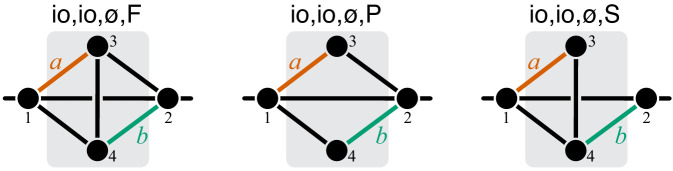
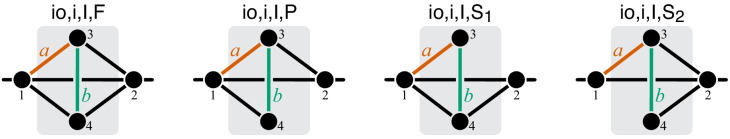
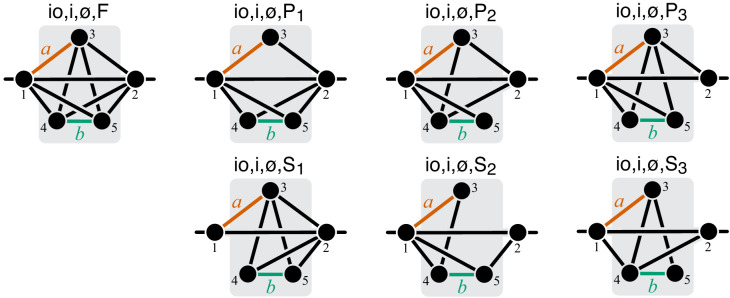
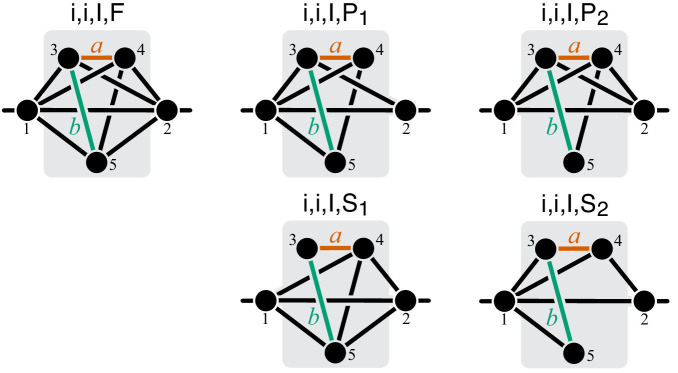
Epistasis is often used to probe functional relationships between genes, and it plays an important role in evolution. However, we lack theory to understand how functional relationships at the molecular level translate into epistasis at the level of whole-organism phenotypes, such as fitness. Here, I derive two rules for how epistasis between mutations with small effects propagates from lower- to higher-level phenotypes in a hierarchical metabolic network with first-order kinetics and how such epistasis depends on topology. Most importantly, weak epistasis at a lower level may be distorted as it propagates to higher levels. Computational analyses show that epistasis in more realistic models likely follows similar, albeit more complex, patterns. These results suggest that pairwise inter-gene epistasis should be common, and it should generically depend on the genetic background and environment. Furthermore, the epistasis coefficients measured for high-level phenotypes may not be sufficient to fully infer the underlying functional relationships.
上位性通常被用来探测基因之间的功能关系,它在进化中起着重要作用。然而,我们缺乏理论来理解分子水平上的功能关系如何转化为整个生物体表型(如适应度)的上位性。在这里,我推导出了在具有一阶动力学的分层代谢网络中,具有小效应的突变之间的上位性如何从较低层次的表型传播到较高层次的表型的两个规则,以及这种上位性如何依赖于拓扑结构。最重要的是,较弱的下位性上位性在传播到较高层次时可能会发生扭曲。计算分析表明,更现实模型中的上位性可能遵循类似的、尽管更复杂的模式。这些结果表明,基因间的成对上位性应该很常见,并且它通常取决于遗传背景和环境。此外,用于高级别表型的上位性系数可能不足以完全推断潜在的功能关系。