Molecular and Cellular Biology Graduate Program, University of Massachusetts, Amherst, United States.
Department of Microbiology, University of Massachusetts, Amherst, United States.
Elife. 2021 Feb 5;10:e60263. doi: 10.7554/eLife.60263.
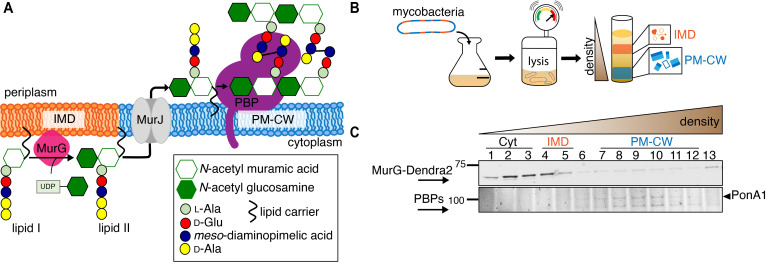
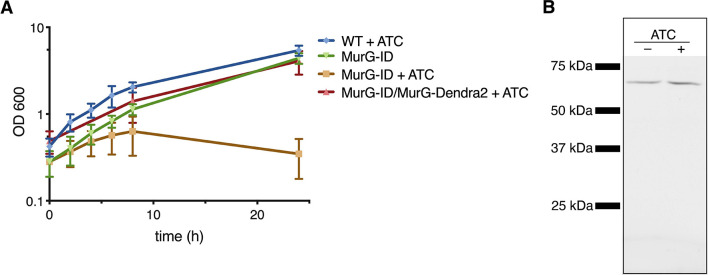
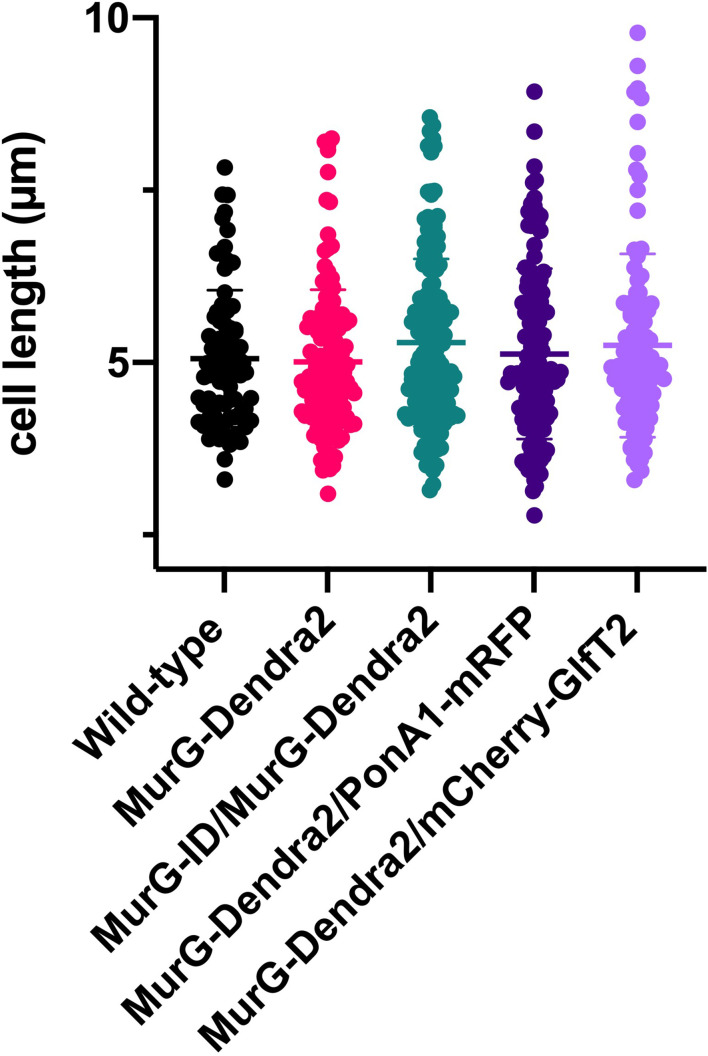
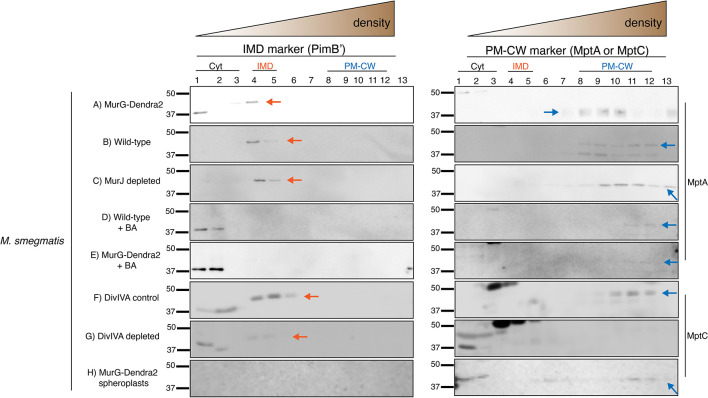
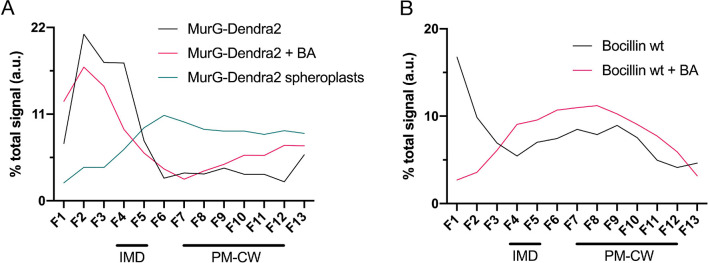
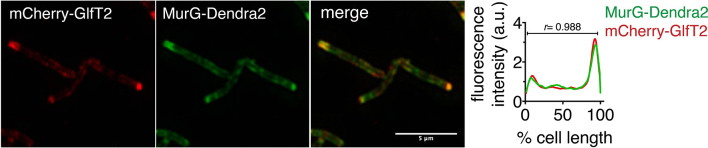
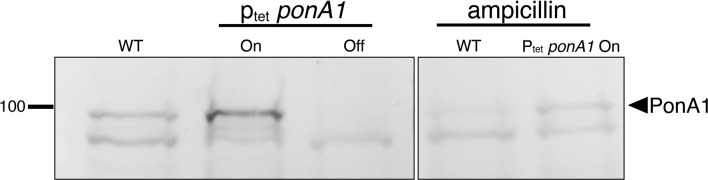
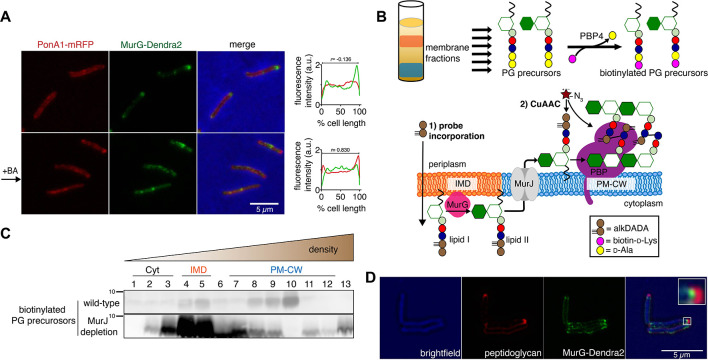
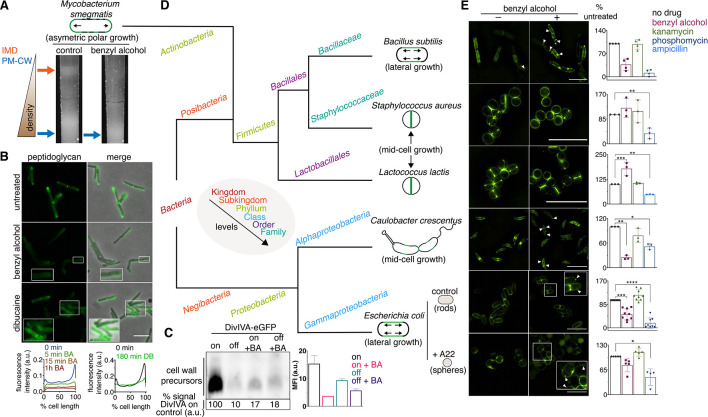
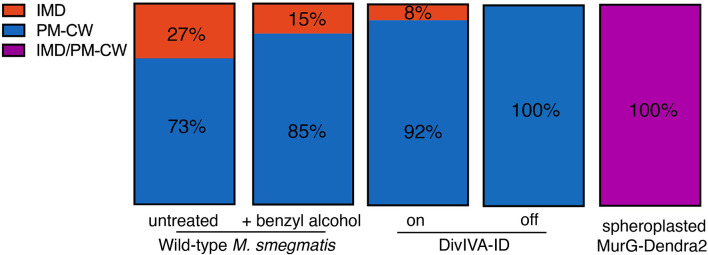
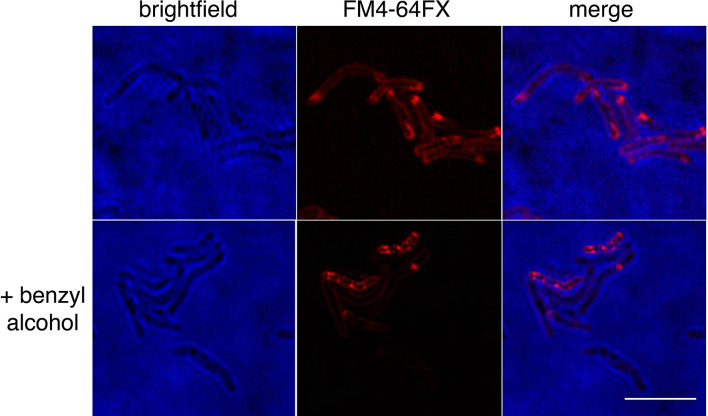
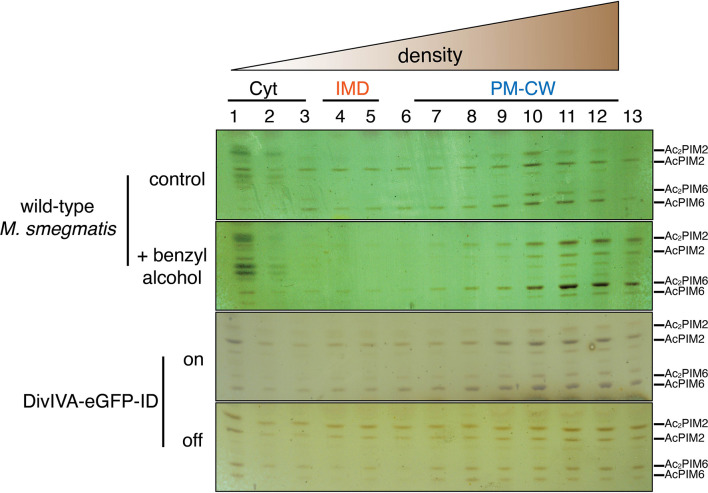
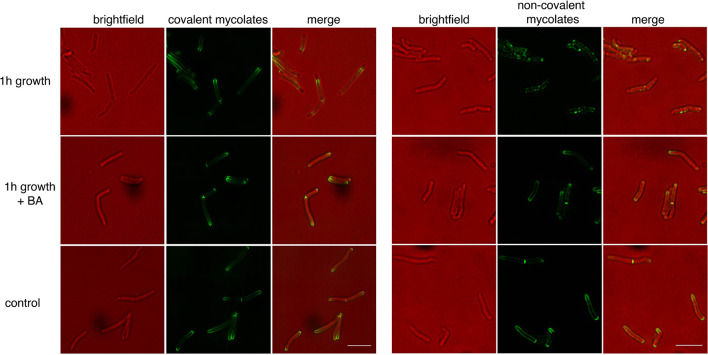
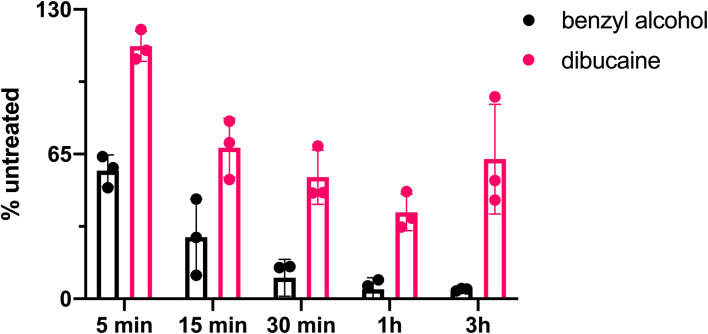
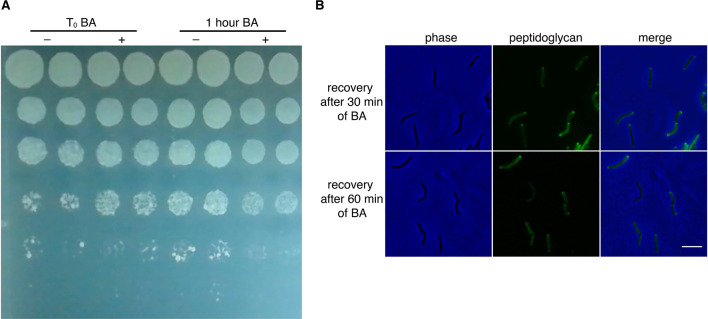
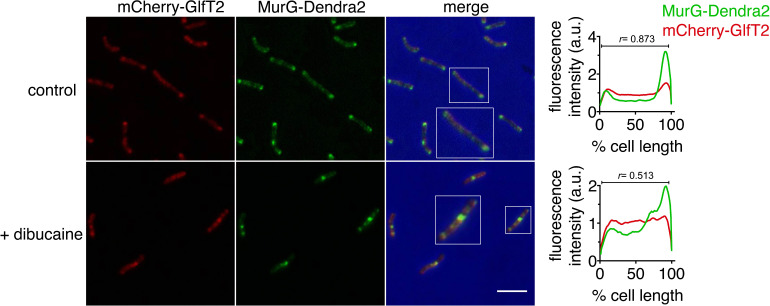
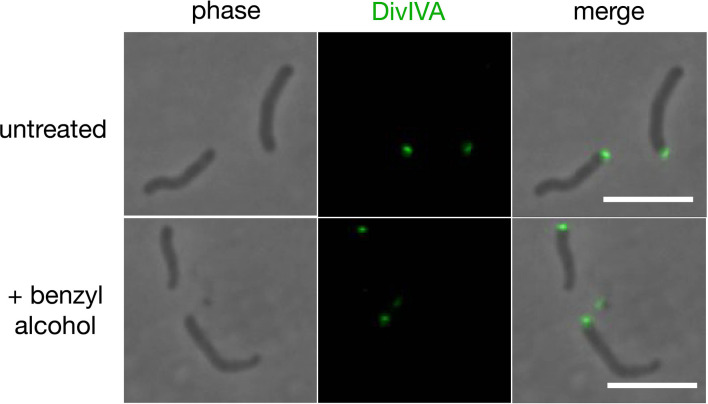
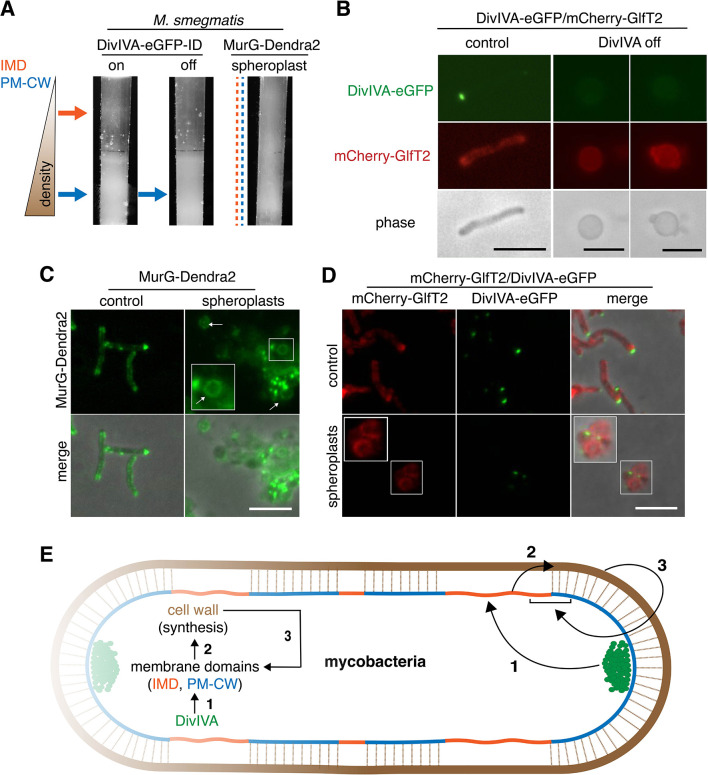
Many antibiotics target the assembly of cell wall peptidoglycan, an essential, heteropolymeric mesh that encases most bacteria. In rod-shaped bacteria, cell wall elongation is spatially precise yet relies on limited pools of lipid-linked precursors that generate and are attracted to membrane disorder. By tracking enzymes, substrates, and products of peptidoglycan biosynthesis in , we show that precursors are made in plasma membrane domains that are laterally and biochemically distinct from sites of cell wall assembly. Membrane partitioning likely contributes to robust, orderly peptidoglycan synthesis, suggesting that these domains help template peptidoglycan synthesis. The cell wall-organizing protein DivIVA and the cell wall itself promote domain homeostasis. These data support a model in which the peptidoglycan polymer feeds back on its membrane template to maintain an environment conducive to directional synthesis. Our findings are applicable to rod-shaped bacteria that are phylogenetically distant from , indicating that horizontal compartmentalization of precursors may be a general feature of bacillary cell wall biogenesis.
许多抗生素针对的是细胞壁肽聚糖的组装,这是一种包裹大多数细菌的必需的、杂多聚的网格。在杆状细菌中,细胞壁的延伸在空间上是精确的,但依赖于有限的脂联肽聚糖前体池,这些前体池产生并被吸引到膜无序区。通过跟踪肽聚糖生物合成中的酶、底物和产物,我们表明前体是在质膜域中产生的,这些域在侧向和生化上与细胞壁组装的部位不同。膜分隔可能有助于稳健、有序的肽聚糖合成,这表明这些域有助于模板肽聚糖合成。细胞壁组织蛋白 DivIVA 和细胞壁本身促进了域内稳态。这些数据支持这样一种模型,即肽聚糖聚合物反馈到其膜模板上,以维持有利于定向合成的环境。我们的发现适用于与 亲缘关系较远的杆状细菌,这表明前体的水平分隔可能是杆状细菌细胞壁生物发生的一个普遍特征。