Institute of Molecular Biology, University of Oregon, Eugene, United States.
Phil and Penny Knight Campus for Accelerating Scientific Impact, University of Oregon, Eugene, United States.
Elife. 2021 Feb 12;10:e64061. doi: 10.7554/eLife.64061.
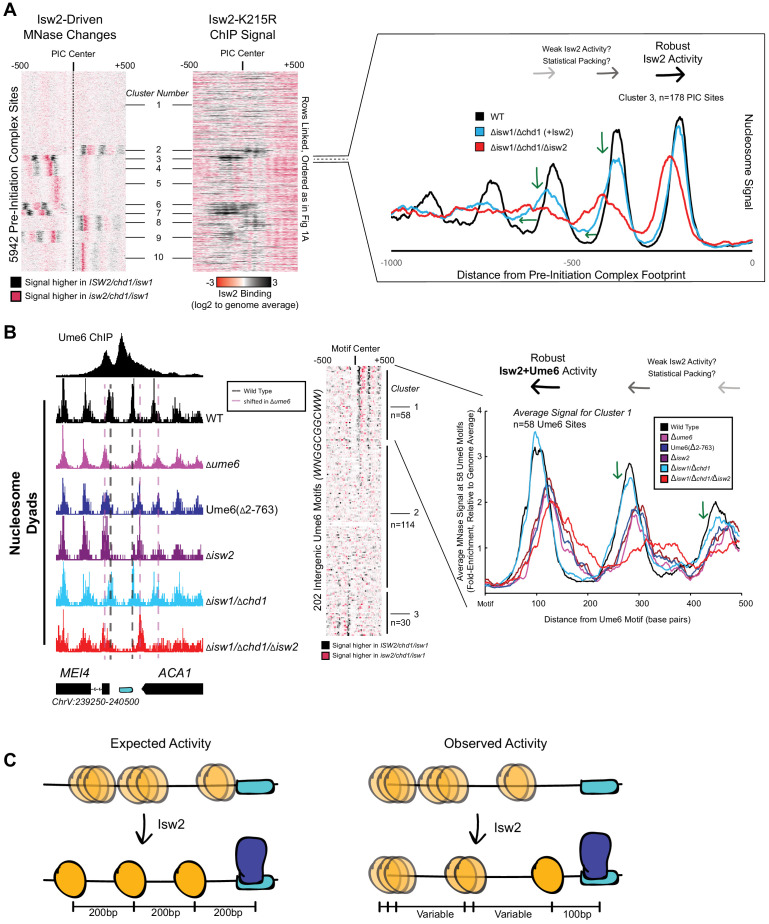
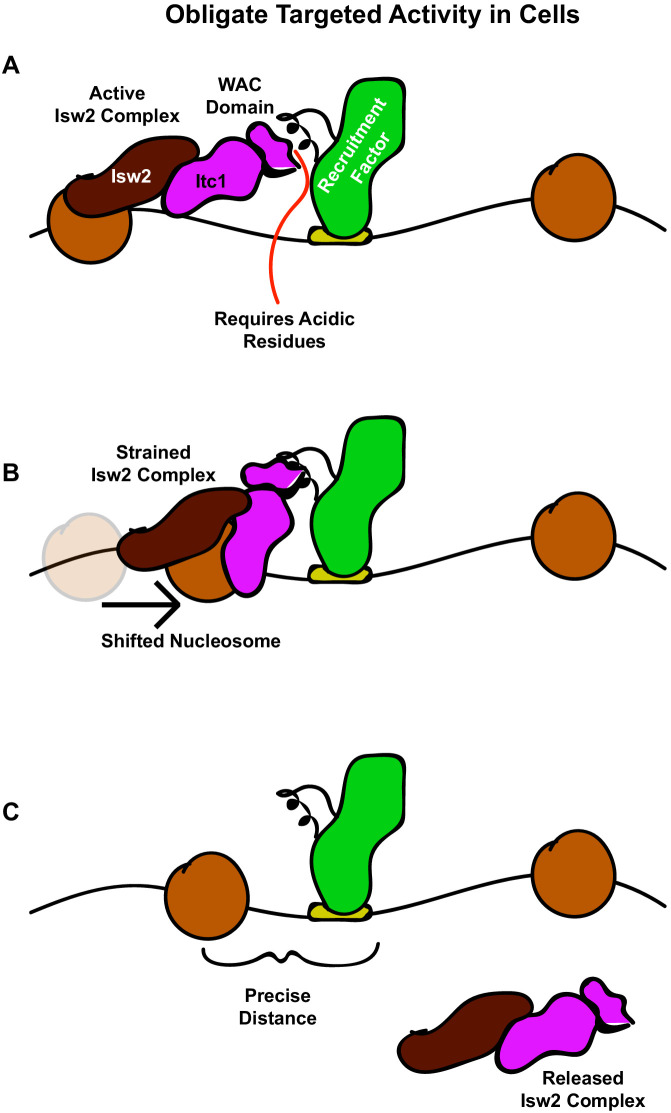
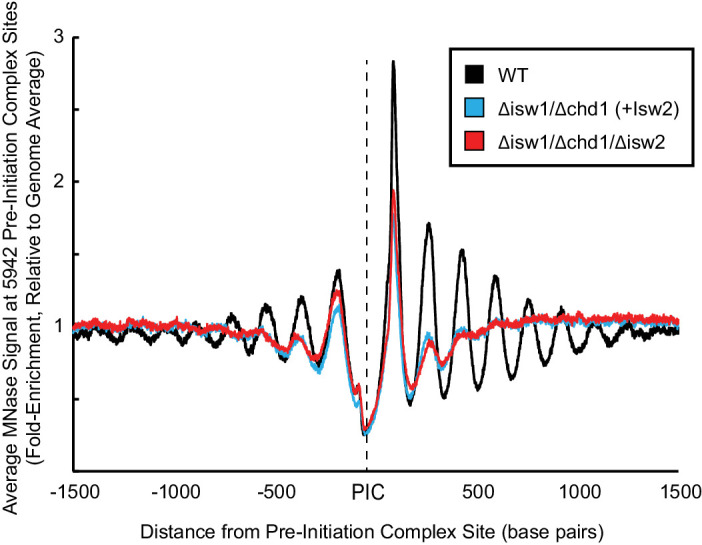
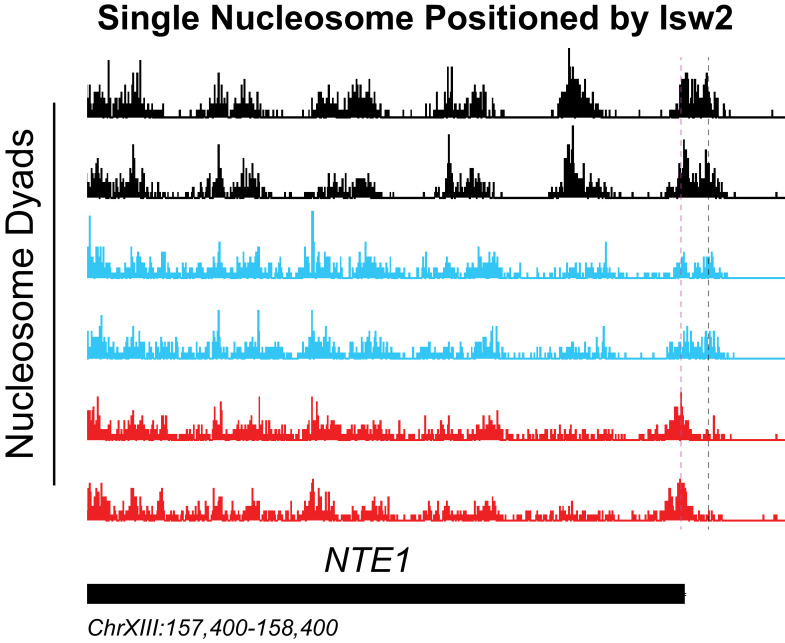
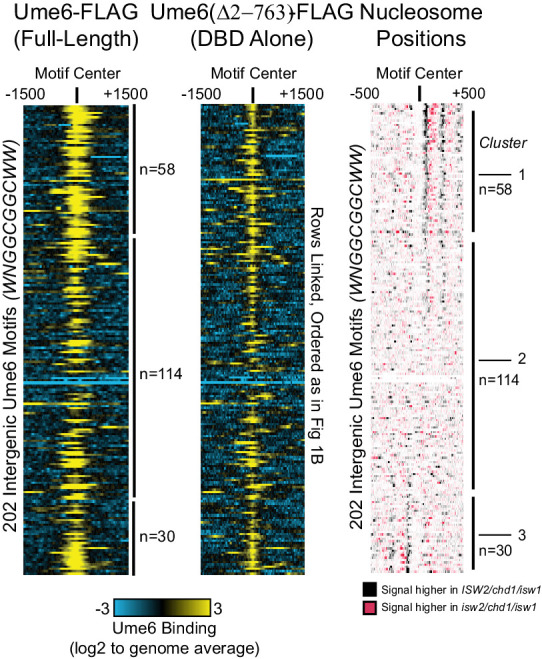
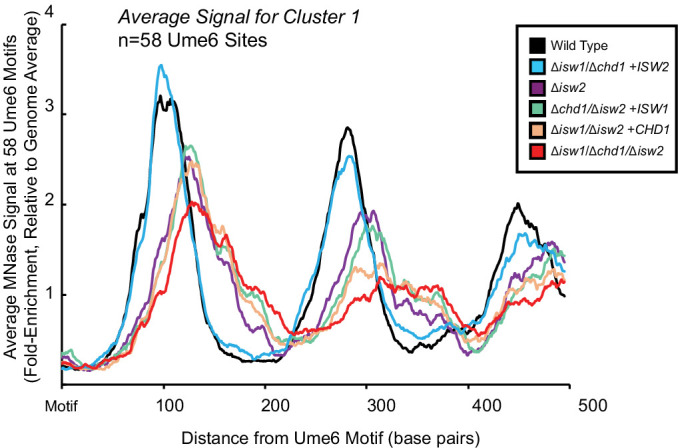
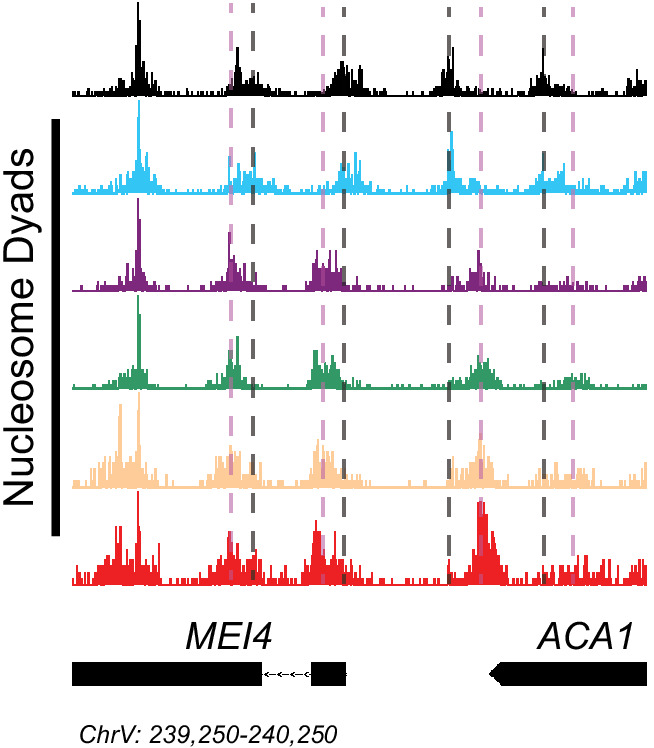
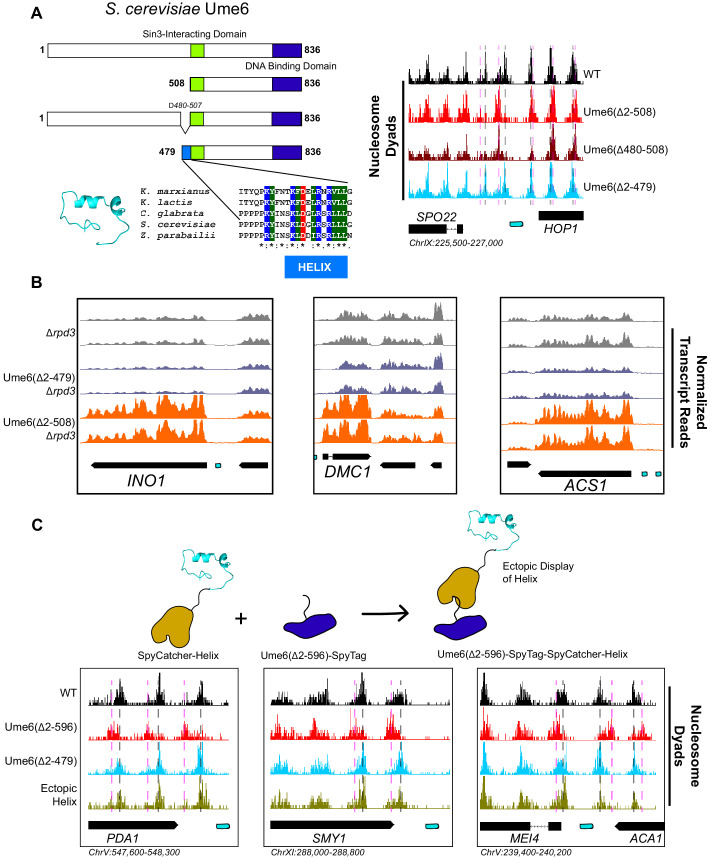
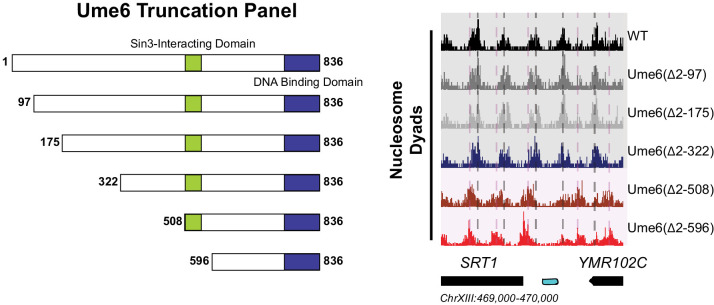
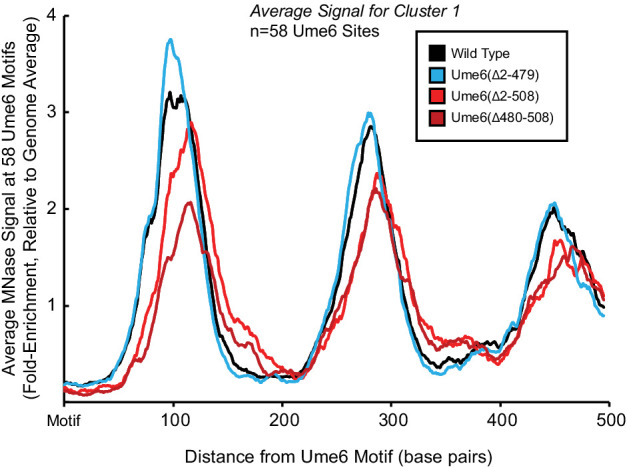
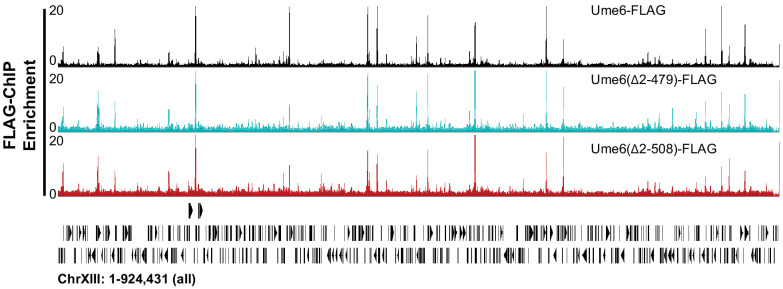
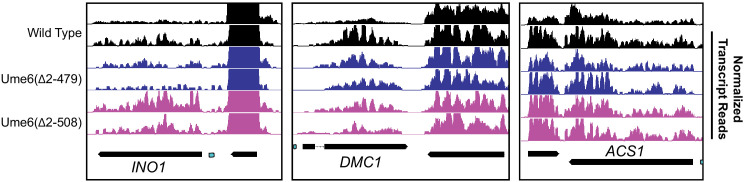
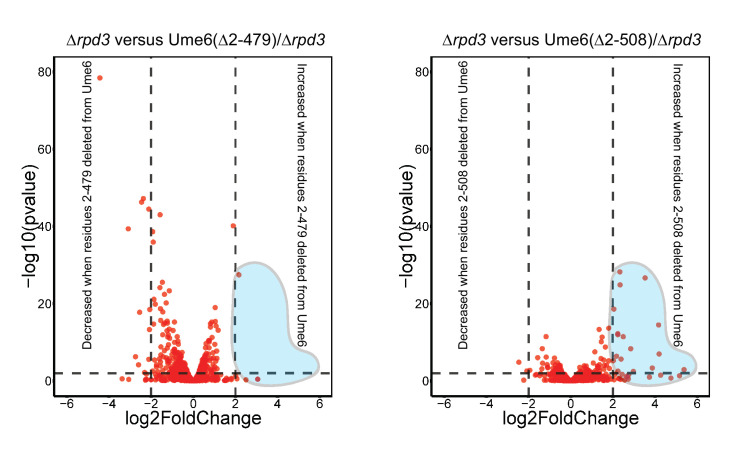
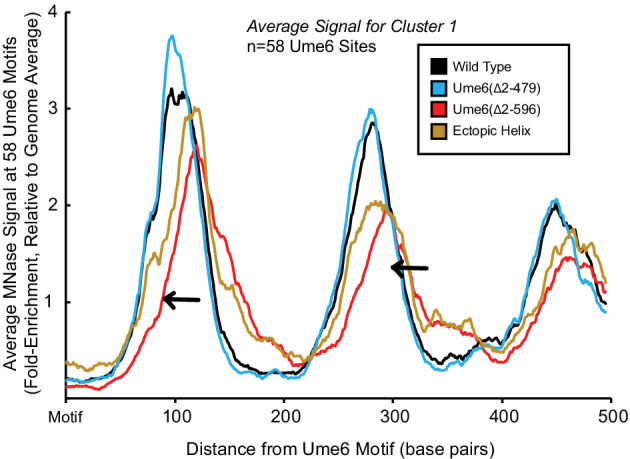
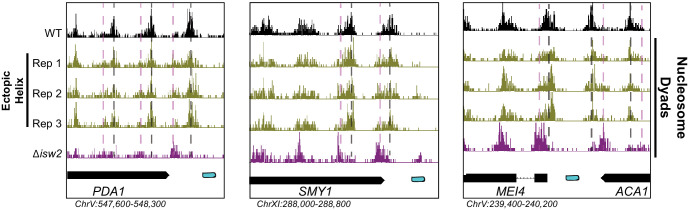
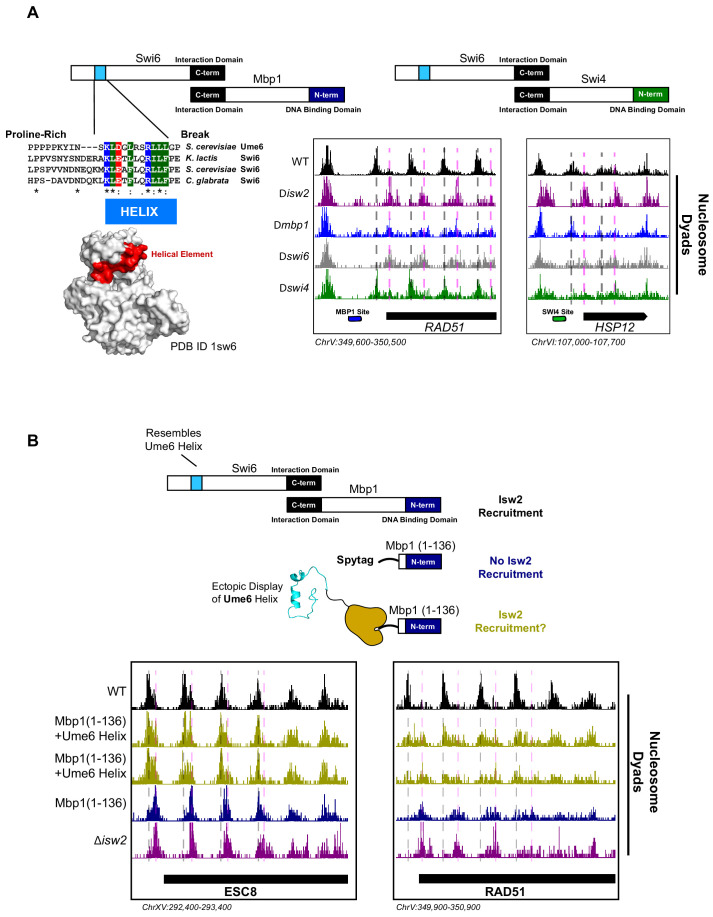
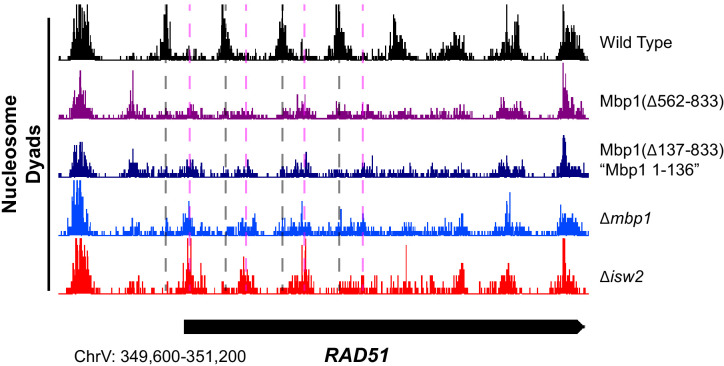
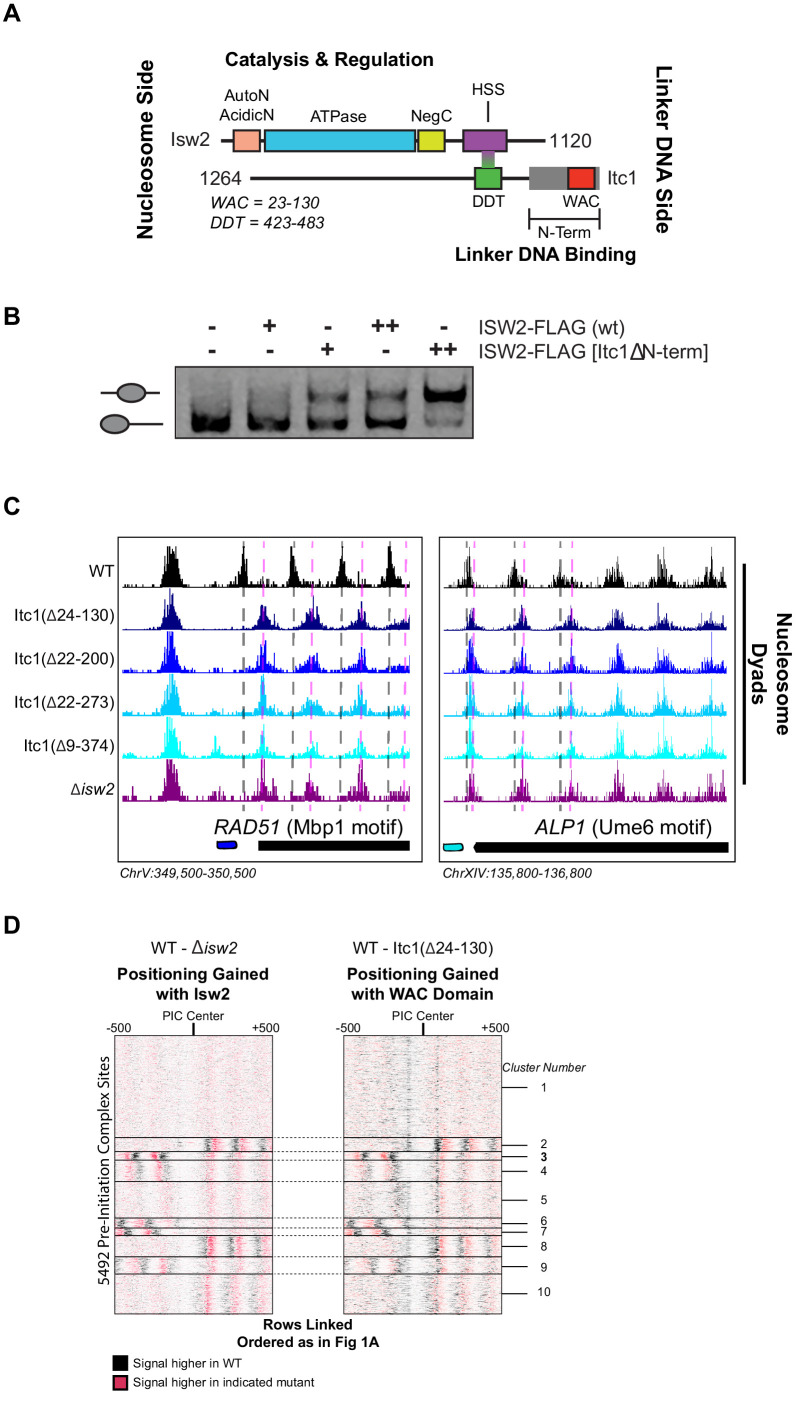
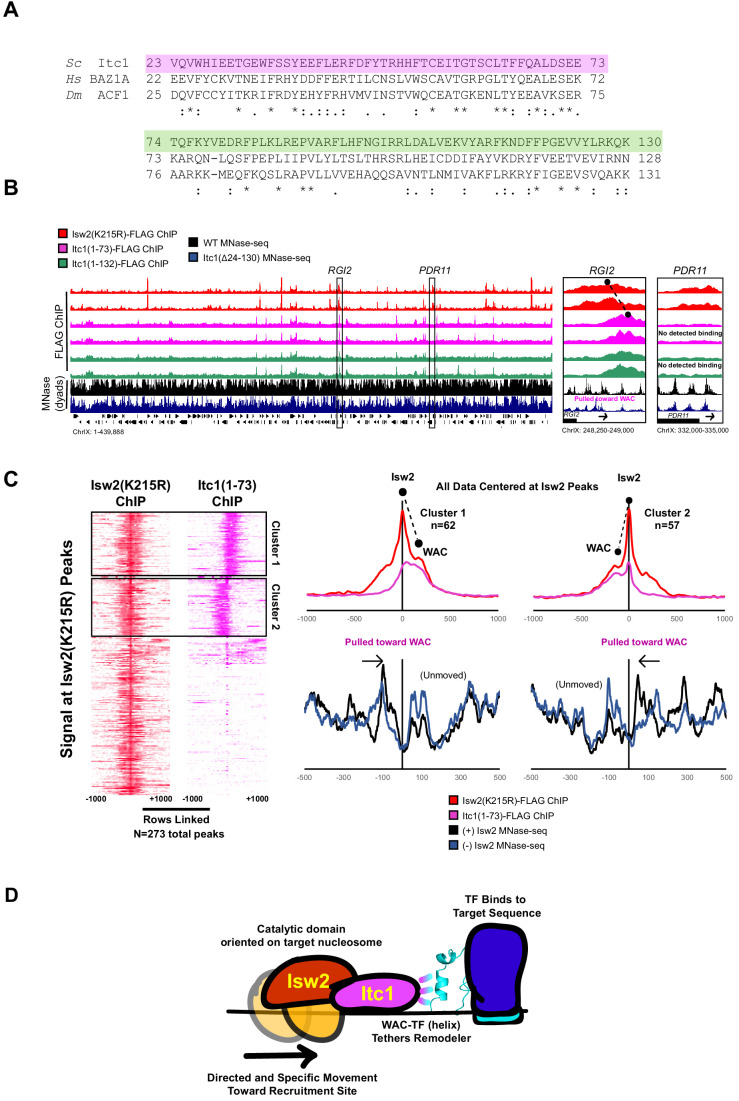
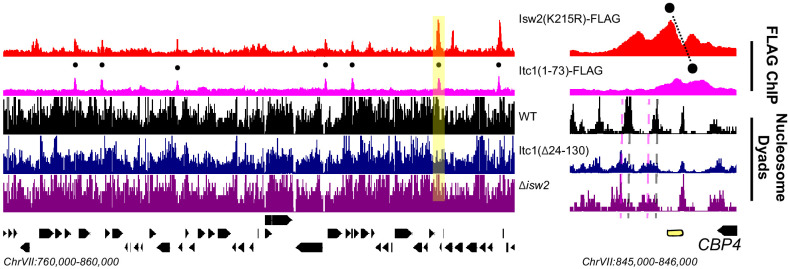
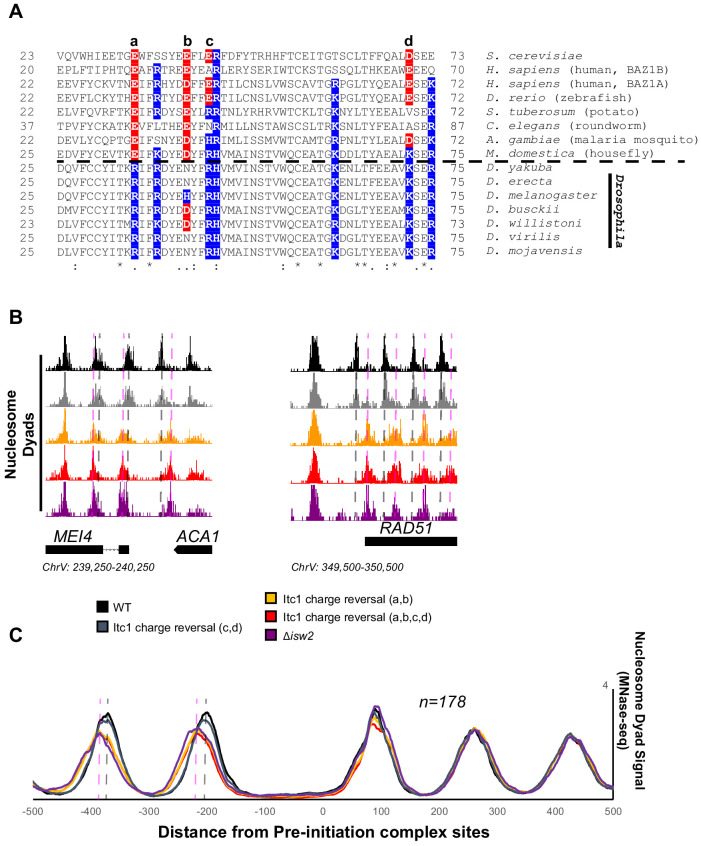
Eukaryotic genomes are organized dynamically through the repositioning of nucleosomes. Isw2 is an enzyme that has been previously defined as a genome-wide, nonspecific nucleosome spacing factor. Here, we show that Isw2 instead acts as an obligately targeted nucleosome remodeler in vivo through physical interactions with sequence-specific factors. We demonstrate that Isw2-recruiting factors use small and previously uncharacterized epitopes, which direct Isw2 activity through highly conserved acidic residues in the Isw2 accessory protein Itc1. This interaction orients Isw2 on target nucleosomes, allowing for precise nucleosome positioning at targeted loci. Finally, we show that these critical acidic residues have been lost in the lineage, potentially explaining the inconsistently characterized function of Isw2-like proteins. Altogether, these data suggest an 'interacting barrier model,' where Isw2 interacts with a sequence-specific factor to accurately and reproducibly position a single, targeted nucleosome to define the precise border of phased chromatin arrays.
真核基因组通过核小体的重定位进行动态组织。Isw2 是一种先前被定义为全基因组非特异性核小体间隔因子的酶。在这里,我们通过与序列特异性因子的物理相互作用表明,Isw2 实际上是一种必需的靶向核小体重塑因子。我们证明,Isw2 募集因子使用小的和以前未表征的表位,这些表位通过 Isw2 辅助蛋白 Itc1 中的高度保守酸性残基引导 Isw2 活性。这种相互作用使 Isw2 定位于靶核小体上,从而在靶位上实现精确的核小体定位。最后,我们表明,这些关键的酸性残基在 谱系中丢失了,这可能解释了 Isw2 样蛋白功能不一致的特征。总之,这些数据表明了一种“相互作用障碍模型”,其中 Isw2 与序列特异性因子相互作用,以准确和可重复地定位单个靶向核小体,从而定义相分离染色质阵列的精确边界。