Princess Margaret Cancer Center, University Health Network, Toronto, Canada.
Princess Margaret Cancer Center, University Health Network, Toronto, Canada; Institute of Medical Science, University of Toronto, Toronto, Canada.
J Biol Chem. 2021 Jan-Jun;296:100505. doi: 10.1016/j.jbc.2021.100505. Epub 2021 Mar 3.
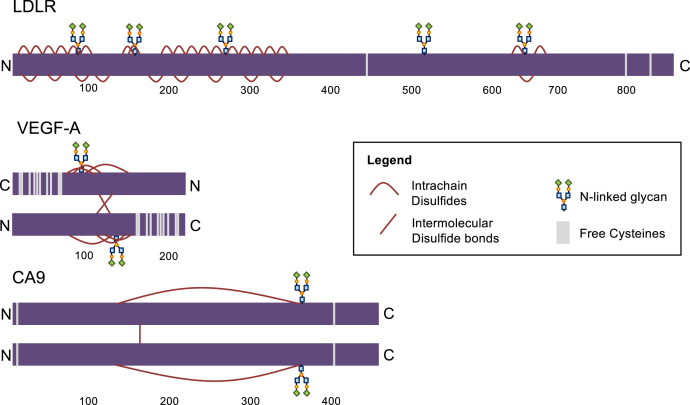
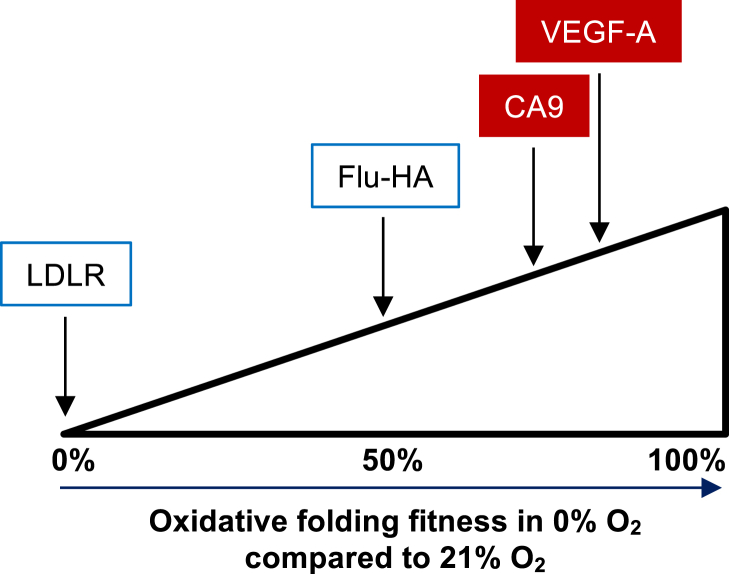
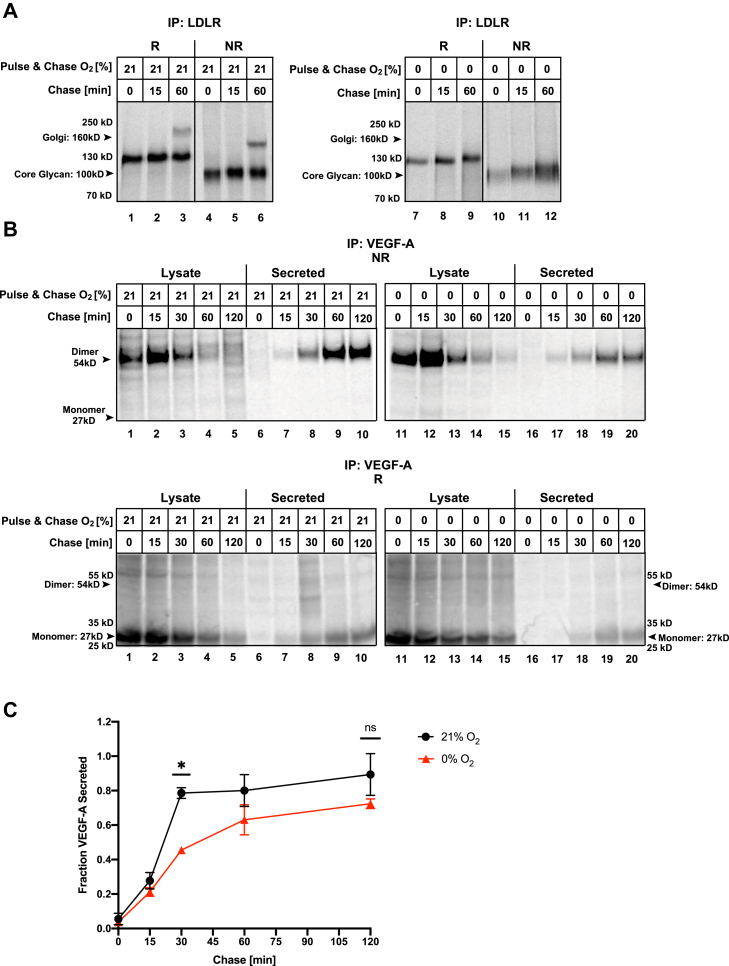
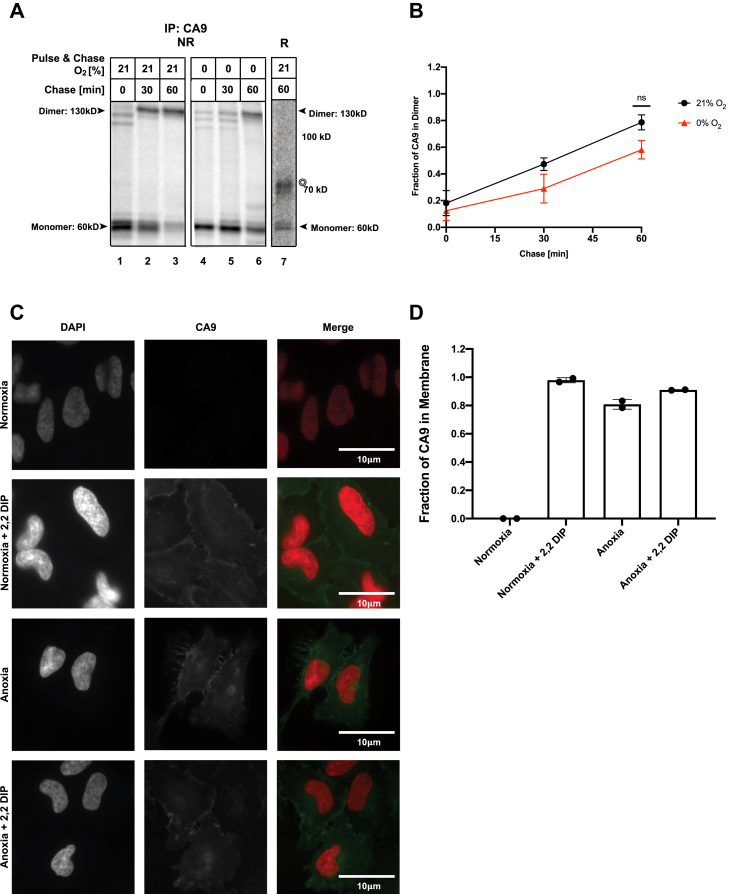
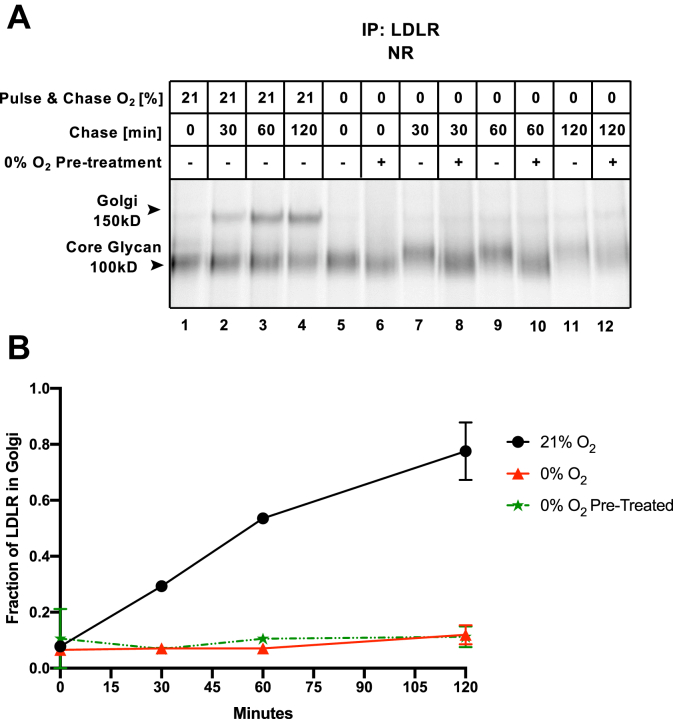
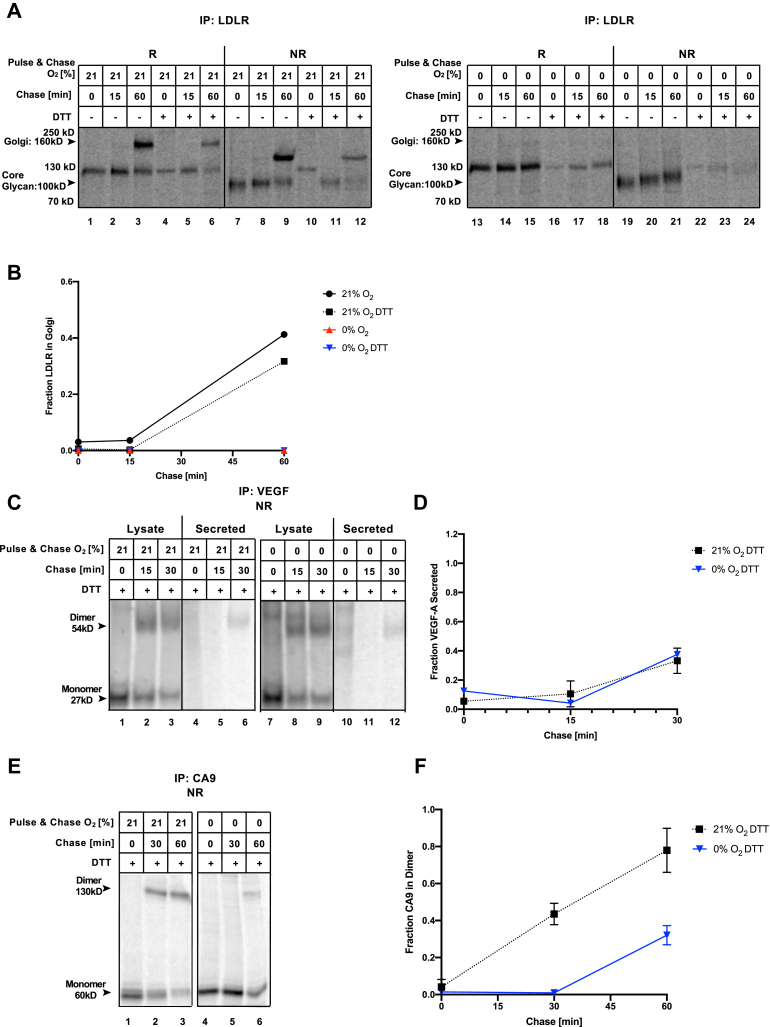
Low levels of oxygen (hypoxia) occurs in many (patho)physiological situations. Adaptation to hypoxia is in part mediated by proteins expressed in the extracellular space that mature in the endoplasmic reticulum (ER) prior to traversing the secretory pathway. The majority of such ER cargo proteins require disulfide bonds for structural stability. Disulfide bonds are formed co- and posttranslationally in a redox relay that requires a terminal electron acceptor such as oxygen. We have previously demonstrated that some ER cargo proteins such as low-density lipoprotein receptor (LDLR) and influenza hemagglutinin (Flu-HA) are unable to complete disulfide bond formation in the absence of oxygen, limiting their ability to pass ER quality control and their ultimate expression. Here, using radioactive pulse-chase immunoprecipitation analysis, we demonstrate that hypoxia-induced ER cargo proteins such as carbonic anhydrase 9 (CA9) and vascular endothelial growth factor A (VEGF-A) complete disulfide bond formation and mature with similar kinetics under hypoxia and normoxia. A global in silico analysis of ER cargo revealed that hypoxia-induced proteins on average contain fewer free cysteines and shorter-range disulfide bonds in comparison to other ER cargo proteins. These data demonstrate the existence of alternative electron acceptors to oxygen for disulfide bond formation in cellulo. However, the ability of different proteins to utilize an oxygen-independent pathway for disulfide bond formation varies widely, contributing to differential gene expression in hypoxia. The superior ability of hypoxia-induced proteins such as VEGF-A and CA9 to mature in hypoxia may be conferred by a simpler disulfide architecture.
低氧(缺氧)发生在许多(病理)生理情况下。缺氧适应部分是通过在穿越分泌途径之前在内质网(ER)中成熟的细胞外空间表达的蛋白质来介导的。大多数这种 ER 货物蛋白需要二硫键来保持结构稳定性。二硫键在需要终端电子受体(如氧气)的氧化还原接力中共同和翻译后形成。我们之前已经证明,一些 ER 货物蛋白,如低密度脂蛋白受体(LDLR)和流感血凝素(Flu-HA),在没有氧气的情况下无法完成二硫键形成,限制了它们通过 ER 质量控制的能力及其最终表达。在这里,我们使用放射性脉冲追踪免疫沉淀分析,证明缺氧诱导的 ER 货物蛋白,如碳酸酐酶 9(CA9)和血管内皮生长因子 A(VEGF-A),在缺氧和常氧条件下完成二硫键形成并以相似的动力学成熟。对 ER 货物的全局计算机分析表明,与其他 ER 货物蛋白相比,缺氧诱导的蛋白质平均含有较少的游离半胱氨酸和较短的二硫键。这些数据表明,在细胞内形成二硫键存在替代氧气的电子受体。然而,不同蛋白质利用非氧依赖性途径形成二硫键的能力差异很大,导致缺氧时基因表达的差异。VEGF-A 和 CA9 等缺氧诱导蛋白在缺氧下成熟的能力更强,这可能归因于更简单的二硫键结构。