Department of Chemistry, Bioscience, and Environmental Engineering, University of Stavanger, Stavanger, Norway.
PLoS One. 2021 Mar 9;16(3):e0241654. doi: 10.1371/journal.pone.0241654. eCollection 2021.
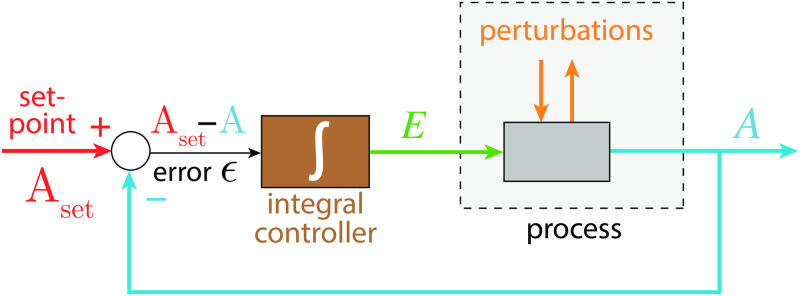
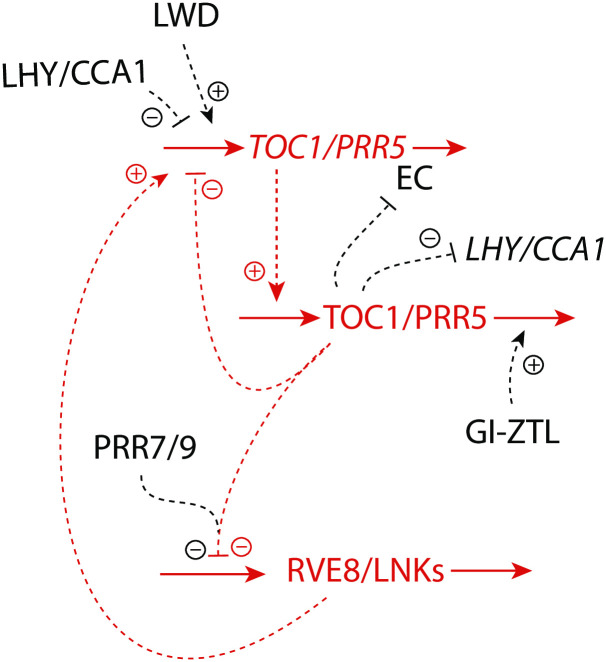
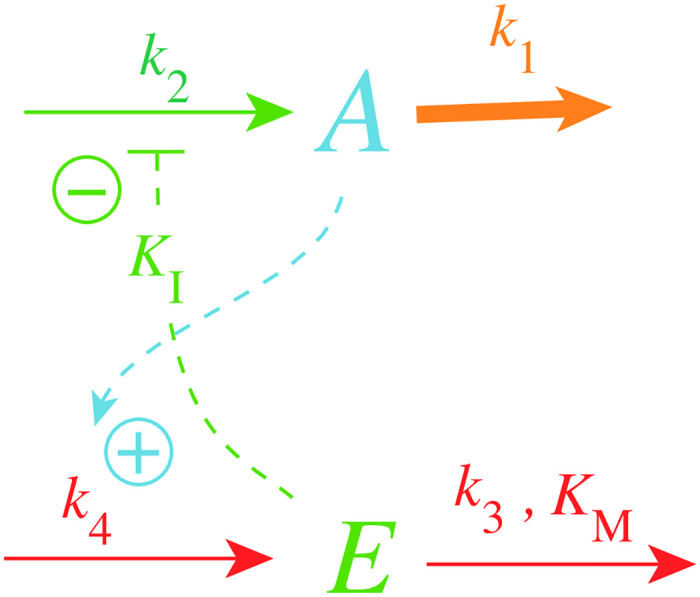
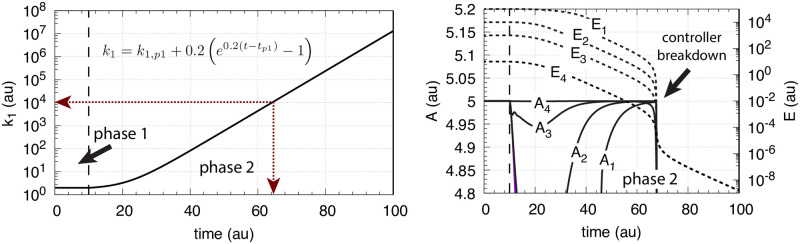
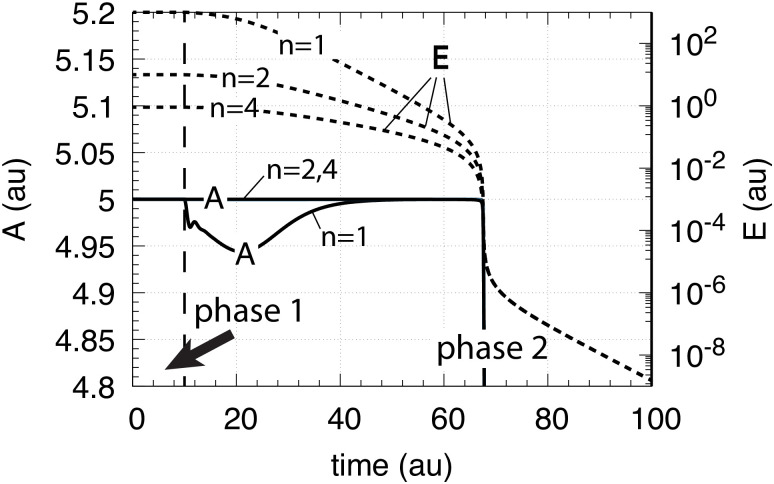
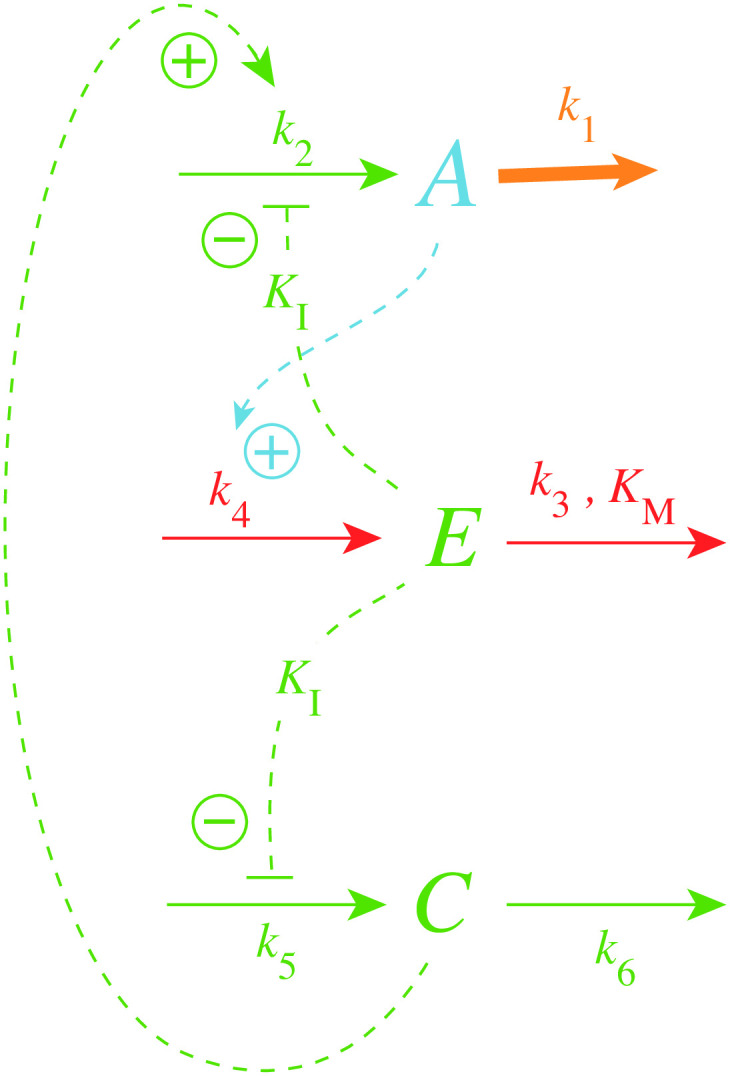
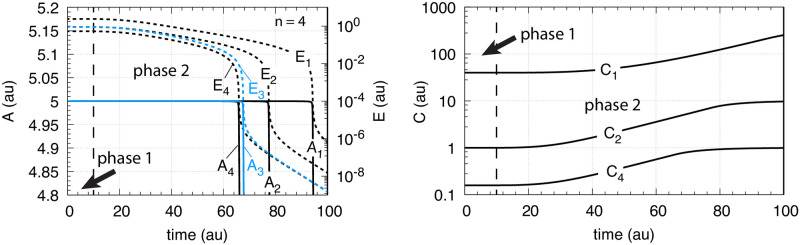
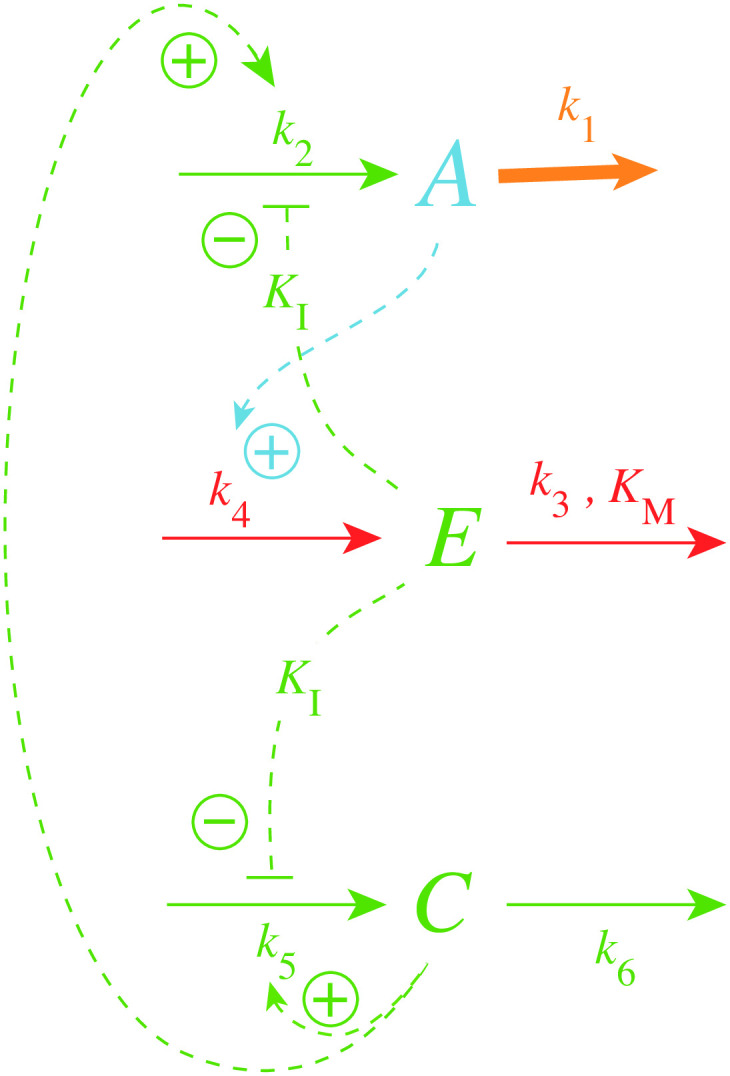
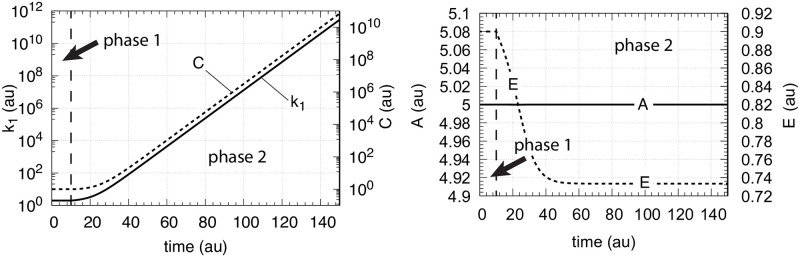
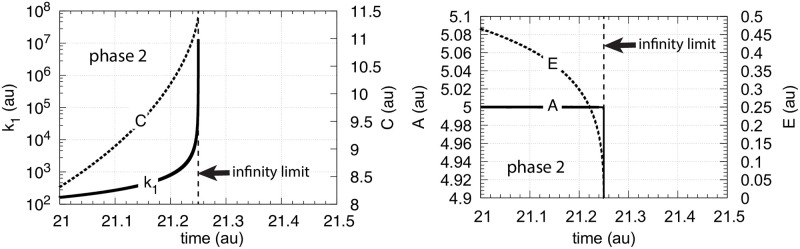
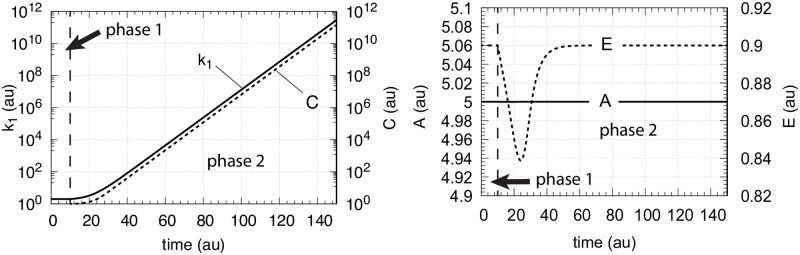
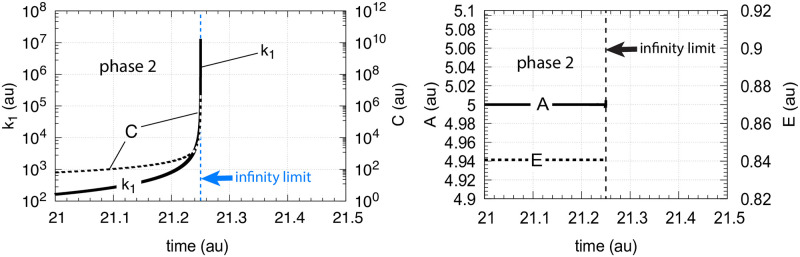
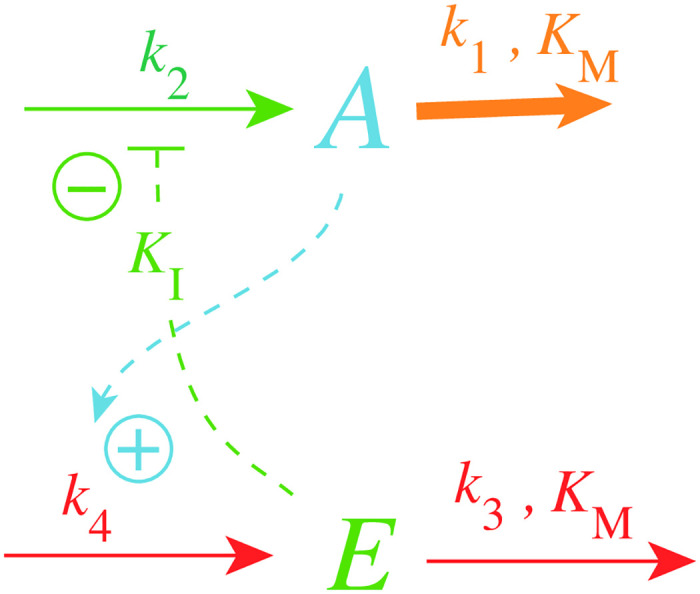
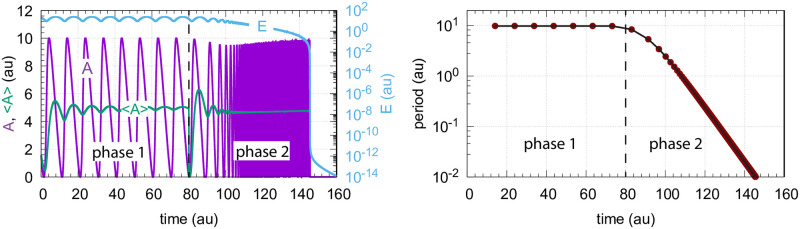
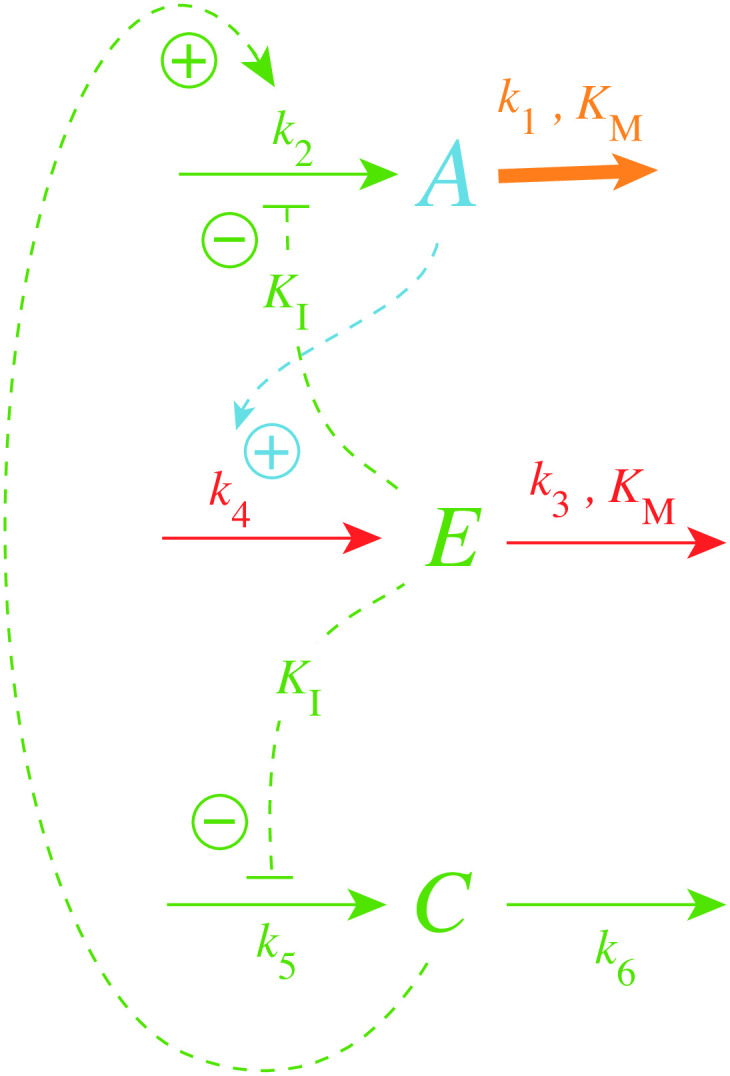
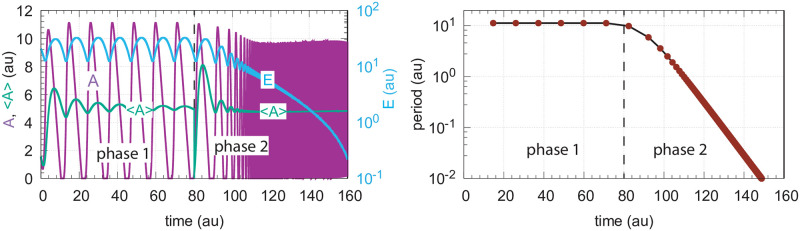
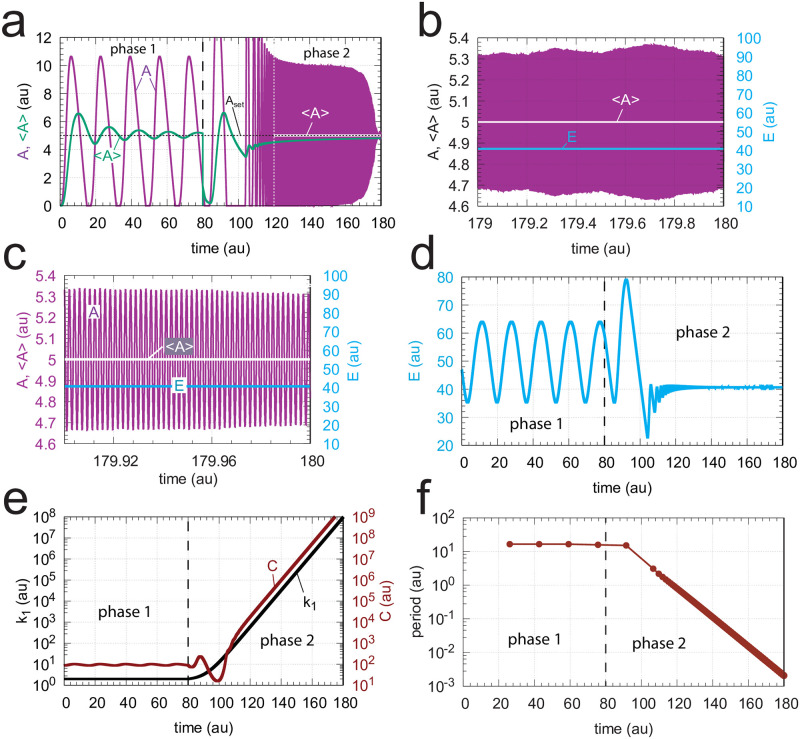
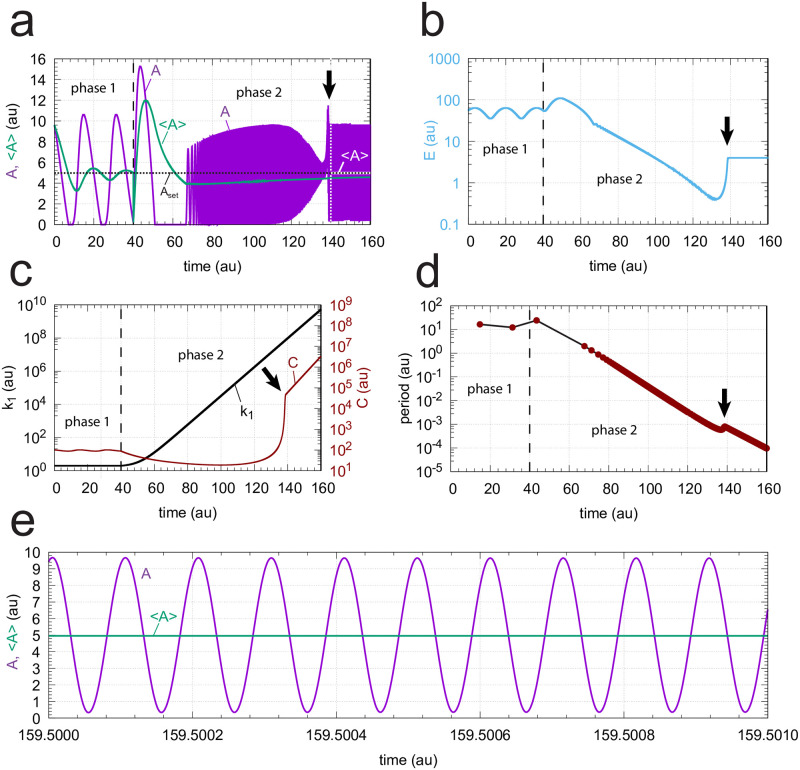
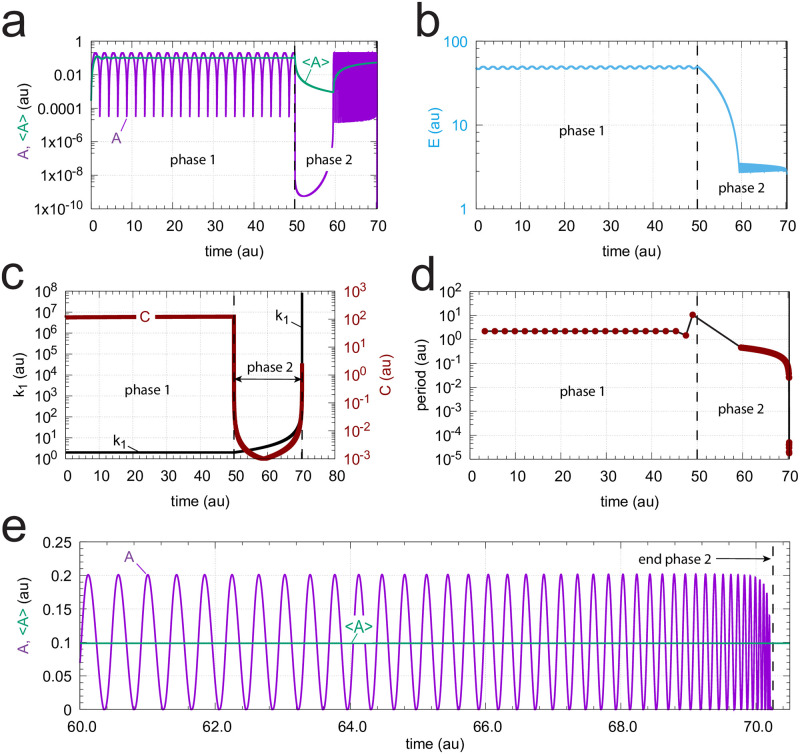
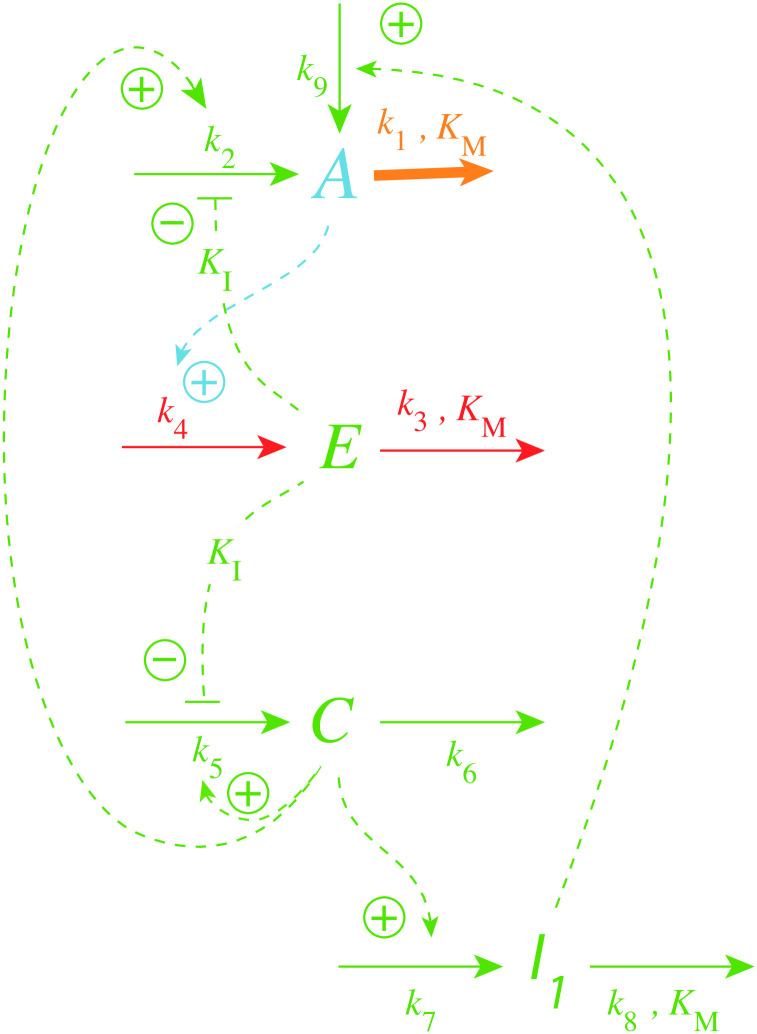
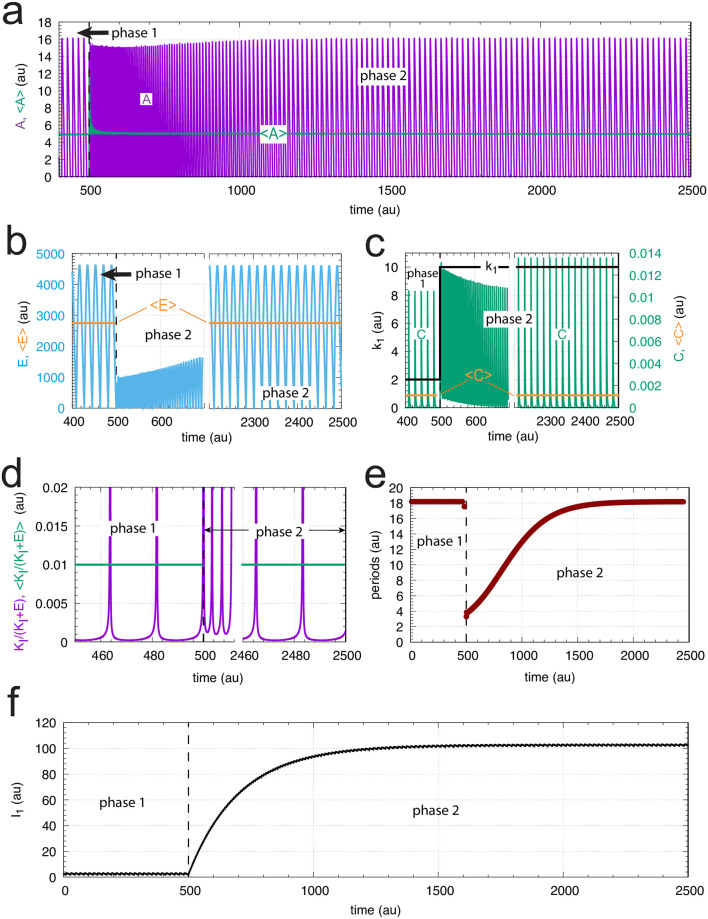
How organisms are able to maintain robust homeostasis has in recent years received increased attention by the use of combined control engineering and kinetic concepts, which led to the discovery of robust controller motifs. While these motifs employ kinetic conditions showing integral feedback and homeostasis for step-wise perturbations, the motifs' performance differ significantly when exposing them to time dependent perturbations. One type of controller motifs which are able to handle exponentially and even hyperbolically growing perturbations are based on derepression. In these controllers the compensatory reaction, which neutralizes the perturbation, is derepressed, i.e. its reaction rate is increased by the decrease of an inhibitor acting on the compensatory flux. While controllers in this category can deal well with different time-dependent perturbations they have the disadvantage that they break down once the concentration of the regulatory inhibitor becomes too low and the compensatory flux has gained its maximum value. We wondered whether it would be possible to bypass this restriction, while still keeping the advantages of derepression kinetics. In this paper we show how the inclusion of multisite inhibition and the presence of positive feedback loops lead to an amplified controller which is still based on derepression kinetics but without showing the breakdown due to low inhibitor concentrations. By searching for the amplified feedback motif in natural systems, we found it as a part of the plant circadian clock where it is highly interlocked with other feedback loops.
近年来,通过将控制工程学和动力学概念相结合的方法,研究生物如何维持强大的体内平衡受到了越来越多的关注,这导致了强大控制器模体的发现。虽然这些模体采用了显示积分反馈和逐步扰动下的体内平衡的动力学条件,但当它们暴露于随时间变化的扰动时,这些模体的性能差异非常显著。一类能够处理指数增长甚至双曲增长的扰动的控制器模体基于去阻遏。在这些控制器中,补偿反应(即中和扰动的反应)被去阻遏,即通过减少作用于补偿通量的抑制剂来增加其反应速率。虽然这类控制器能够很好地处理不同的时变扰动,但它们有一个缺点,即一旦调节抑制剂的浓度变得过低,补偿通量达到最大值,控制器就会失效。我们想知道是否有可能绕过这个限制,同时仍然保留去阻遏动力学的优势。在本文中,我们展示了如何通过包含多部位抑制和存在正反馈回路来放大控制器,从而仍然基于去阻遏动力学,但不会因抑制剂浓度低而失效。通过在自然系统中寻找放大的反馈模体,我们发现它是植物生物钟的一部分,它与其他反馈回路高度交织在一起。