Department of Biochemistry and Biophysics, University of California at San Francisco, San Francisco, United States.
Department of Biochemistry, University of Washington, Seattle, United States.
Elife. 2021 May 5;10:e65168. doi: 10.7554/eLife.65168.
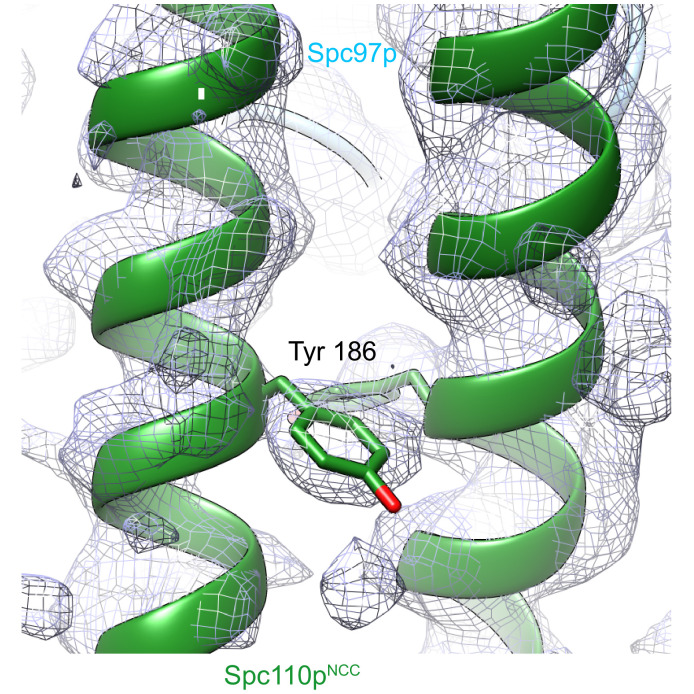
Microtubule (MT) nucleation is regulated by the γ-tubulin ring complex (γTuRC), conserved from yeast to humans. In , γTuRC is composed of seven identical γ-tubulin small complex (γTuSC) sub-assemblies, which associate helically to template MT growth. γTuRC assembly provides a key point of regulation for the MT cytoskeleton. Here, we combine crosslinking mass spectrometry, X-ray crystallography, and cryo-EM structures of both monomeric and dimeric γTuSCs, and open and closed helical γTuRC assemblies in complex with Spc110p to elucidate the mechanisms of γTuRC assembly. γTuRC assembly is substantially aided by the evolutionarily conserved CM1 motif in Spc110p spanning a pair of adjacent γTuSCs. By providing the highest resolution and most complete views of any γTuSC assembly, our structures allow phosphorylation sites to be mapped, surprisingly suggesting that they are mostly inhibitory. A comparison of our structures with the CM1 binding site in the human γTuRC structure at the interface between GCP2 and GCP6 allows for the interpretation of significant structural changes arising from CM1 helix binding to metazoan γTuRC.
微管 (MT) 的成核受 γ-微管蛋白环复合物 (γTuRC) 的调控,从酵母到人都有这种复合物。在 ,γTuRC 由七个相同的 γ-微管蛋白小复合物 (γTuSC) 亚基组装体组成,这些亚基以螺旋方式组装,为 MT 的生长提供模板。γTuRC 的组装为 MT 细胞骨架的调控提供了一个关键点。在这里,我们结合交联质谱、X 射线晶体学和单体及二聚体 γTuSC 以及与 Spc110p 结合的开放和闭合螺旋 γTuRC 组装体的 cryo-EM 结构,阐明了 γTuRC 组装的机制。γTuRC 的组装在 Spc110p 中的进化保守的 CM1 基序的帮助下大大增强,该基序跨越一对相邻的 γTuSCs。通过提供任何 γTuSC 组装体的最高分辨率和最完整的视图,我们的结构允许对磷酸化位点进行映射,令人惊讶的是,这些位点大多是抑制性的。我们的结构与人类 γTuRC 结构中 GCP2 和 GCP6 之间界面处的 CM1 结合位点的比较允许解释由于 CM1 螺旋结合到后生动物 γTuRC 而产生的显著结构变化。