Department of Anatomy and Cell Biology, George Washington University School of Medicine and Health Sciences, Washington, United States.
GW Cancer Center, George Washington University School of Medicine and Health Sciences, Washington, United States.
Elife. 2021 May 18;10:e65770. doi: 10.7554/eLife.65770.
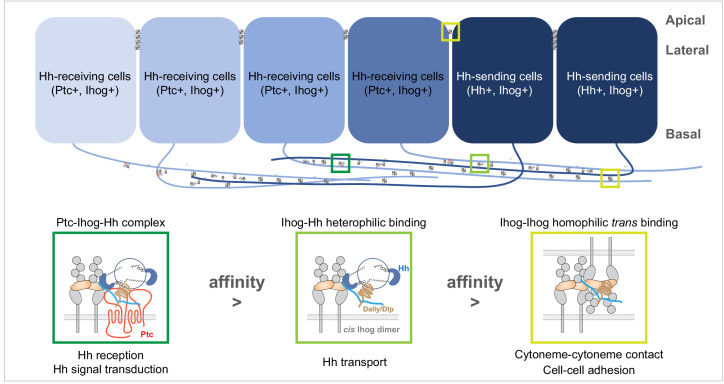
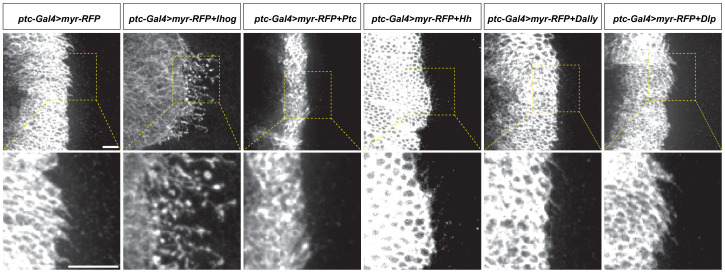
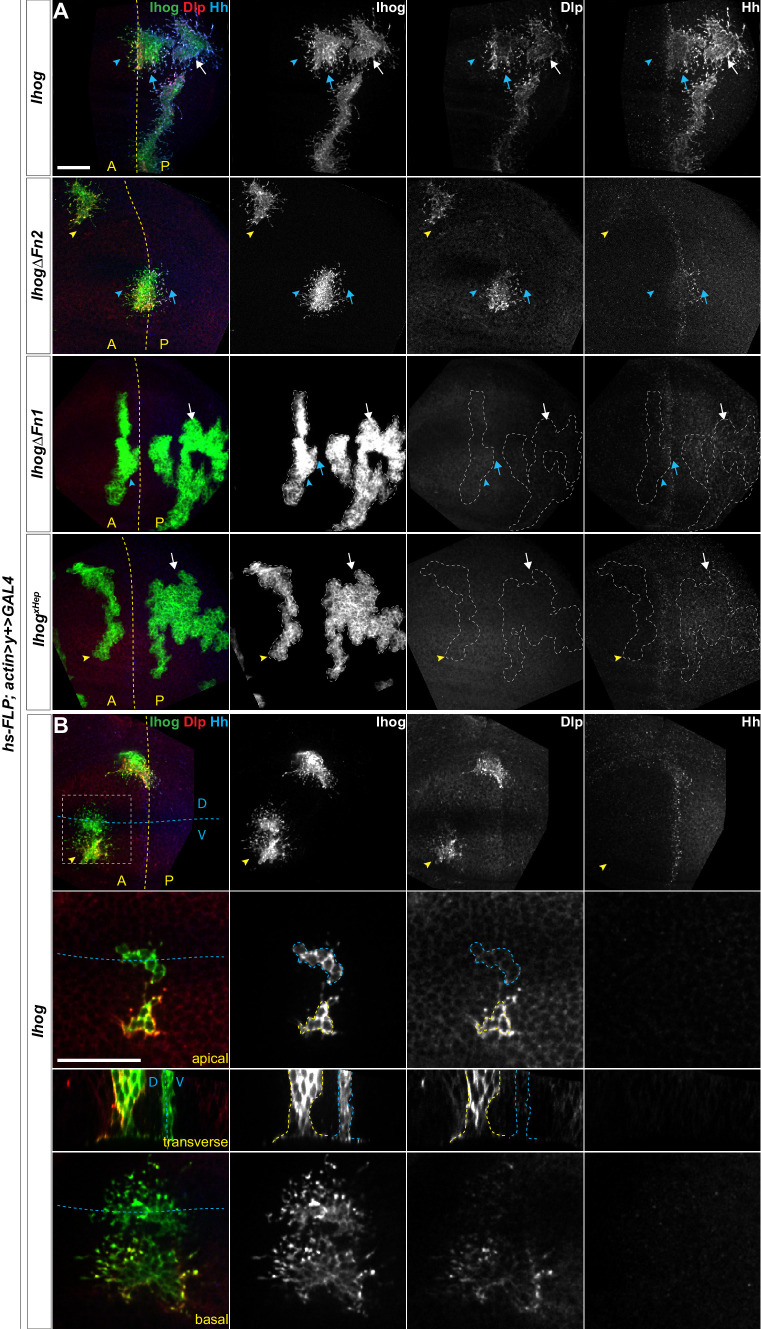
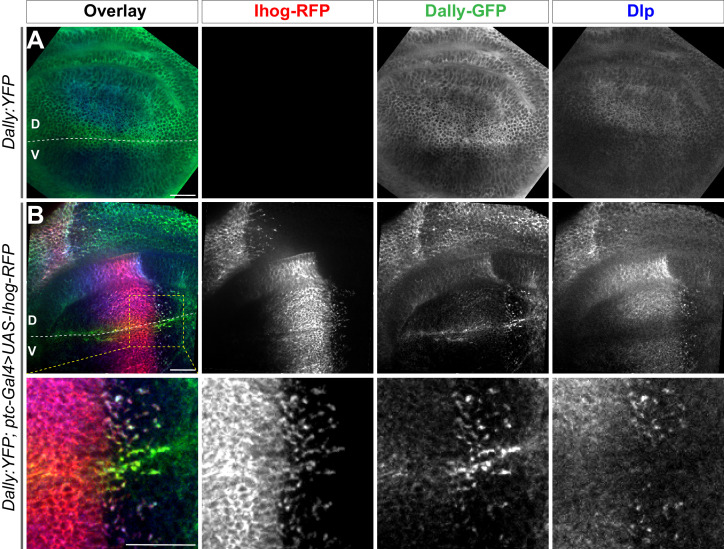
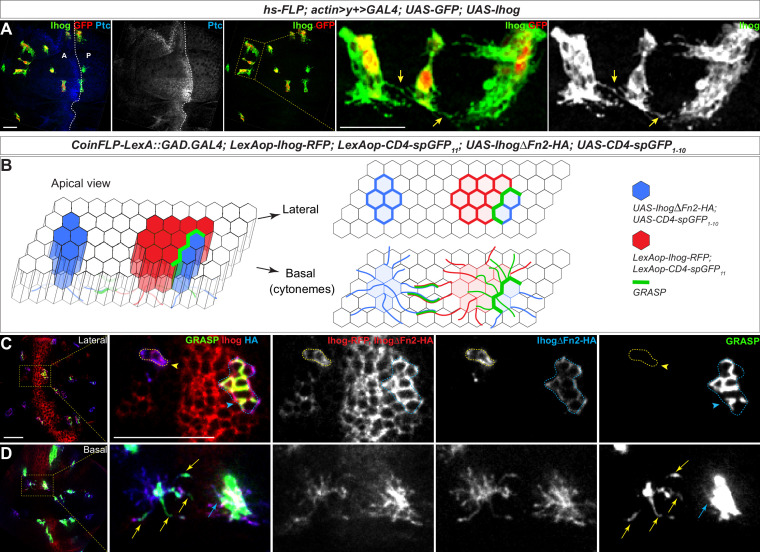
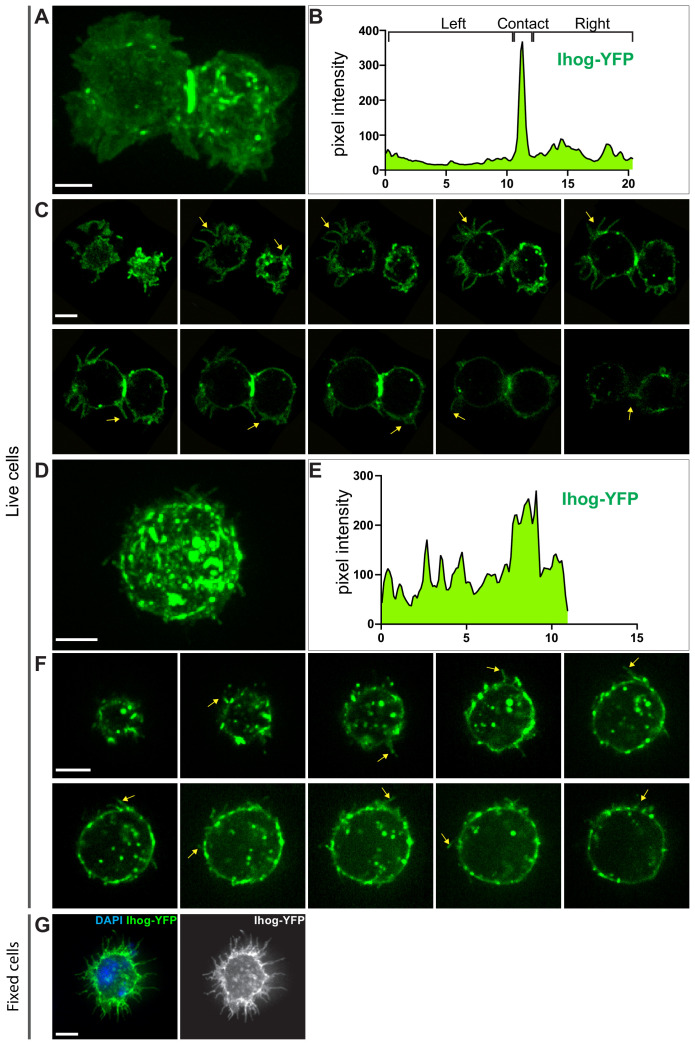
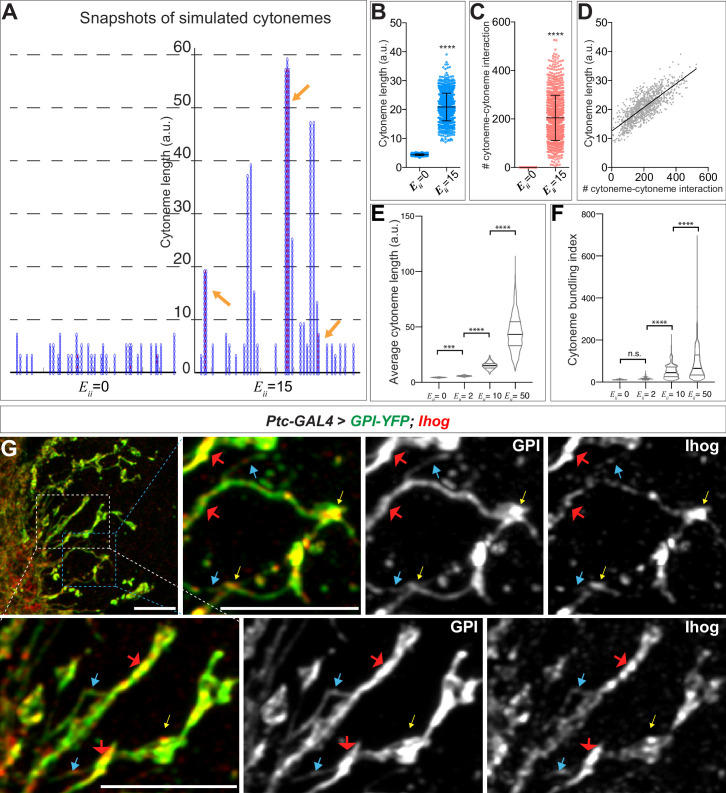
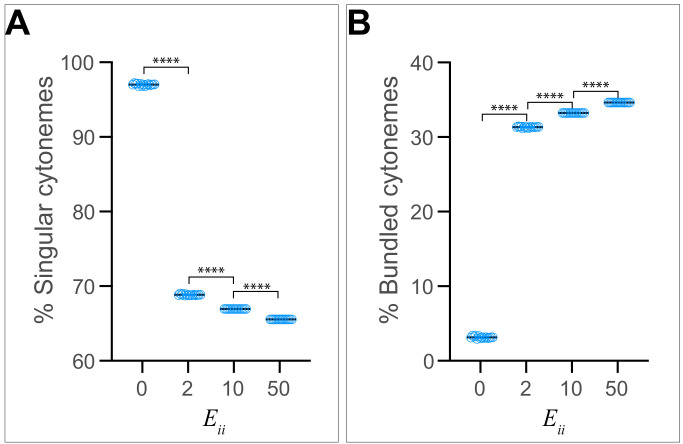
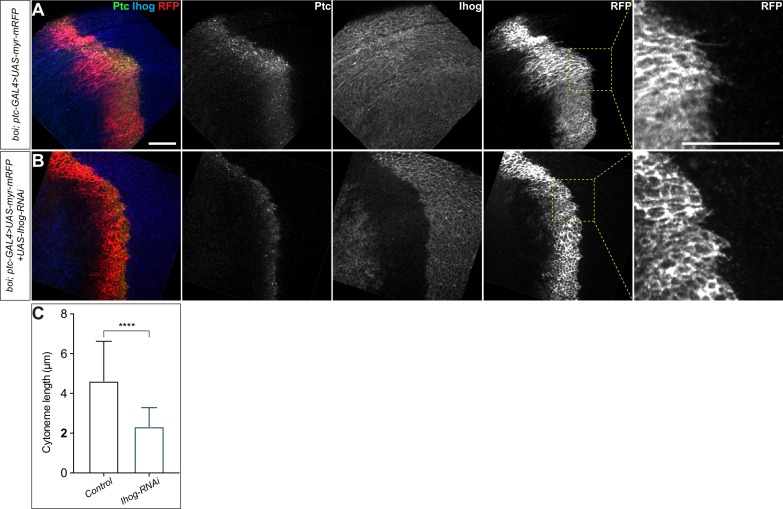
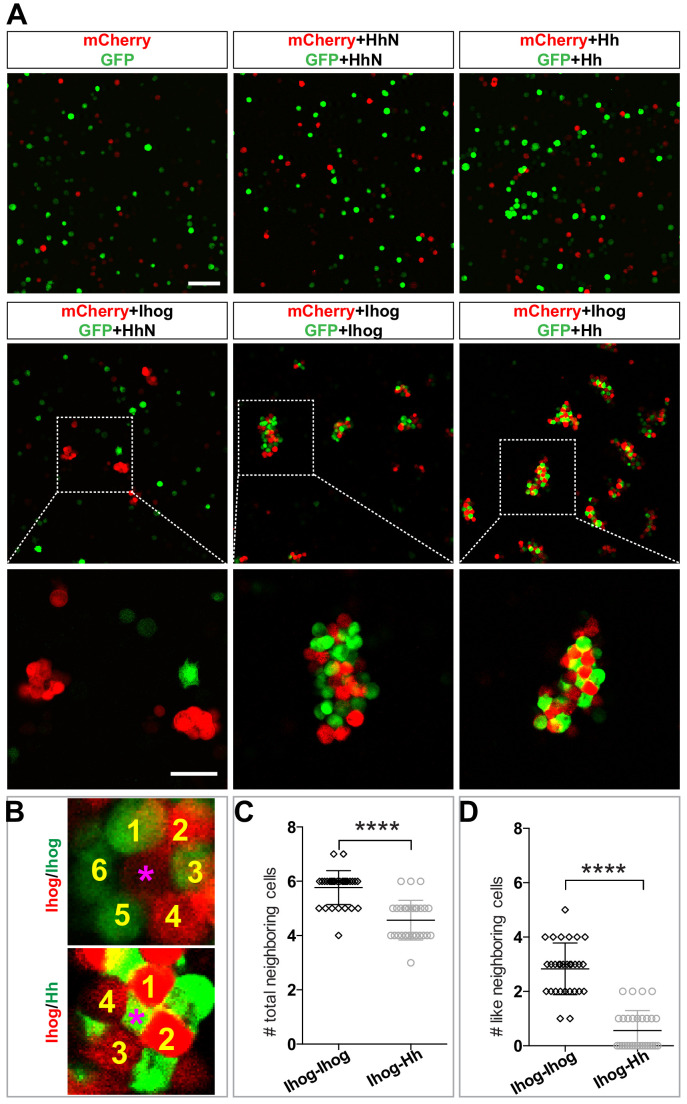
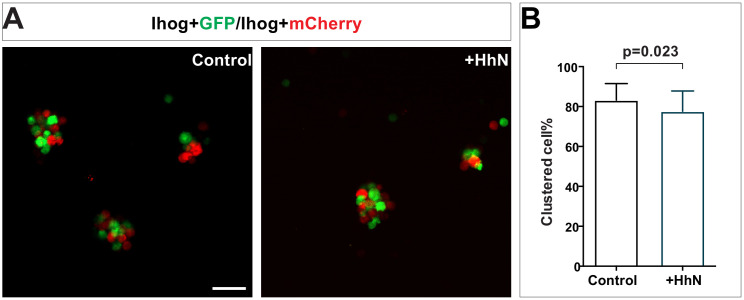
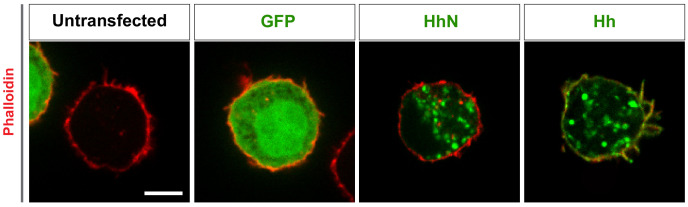
Hedgehog (Hh) signaling patterns embryonic tissues and contributes to homeostasis in adults. In , Hh transport and signaling are thought to occur along a specialized class of actin-rich filopodia, termed cytonemes. Here, we report that Interference hedgehog (Ihog) not only forms a Hh receptor complex with Patched to mediate intracellular signaling, but Ihog also engages in -homophilic binding leading to cytoneme stabilization in a manner independent of its role as the Hh receptor. Both functions of Ihog (-homophilic binding for cytoneme stabilization and Hh binding for ligand sensing) involve a heparin-binding site on the first fibronectin repeat of the extracellular domain. Thus, the Ihog-Ihog interaction and the Hh-Ihog interaction cannot occur simultaneously for a single Ihog molecule. By combining experimental data and mathematical modeling, we determined that Hh-Ihog heterophilic interaction dominates and Hh can disrupt and displace Ihog molecules involved in -homophilic binding. Consequently, we proposed that the weaker Ihog-Ihog interaction promotes and stabilizes direct membrane contacts along cytonemes and that, as the cytoneme encounters secreted Hh ligands, the ligands trigger release of Ihog from Ihog-Ihog complex enabling transport or internalization of the Hh ligand-Ihog-Patched -receptor complex. Thus, the seemingly incompatible functions of Ihog in homophilic adhesion and ligand binding cooperate to assist Hh transport and reception along the cytonemes.
刺猬(Hh)信号模式胚胎组织,并有助于在成人的体内平衡。在,Hh 运输和信号被认为沿着一个特殊的肌动蛋白丰富的纤毛,称为纤毛发生。在这里,我们报告说干扰刺猬(Ihog)不仅与帕奇特(Patched)形成 Hh 受体复合物来介导细胞内信号转导,而且 Ihog 还参与同种型结合,导致纤毛发生稳定,其方式独立于其作为 Hh 受体的作用。Ihog 的这两种功能(同种型结合以稳定纤毛发生和结合 Hh 以感知配体)都涉及细胞外结构域第一纤维连接蛋白重复中的肝素结合位点。因此,单个 Ihog 分子不能同时发生 Ihog-Ihog 相互作用和 Hh-Ihog 相互作用。通过结合实验数据和数学模型,我们确定 Hh-Ihog 异源相互作用占主导地位,并且 Hh 可以破坏和取代参与同种型结合的 Ihog 分子。因此,我们提出较弱的 Ihog-Ihog 相互作用促进并稳定了纤毛发生中的直接膜接触,并且当纤毛发生遇到分泌的 Hh 配体时,配体触发 Ihog 从 Ihog-Ihog 复合物中释放出来,从而使 Hh 配体-Ihog-Patched 受体复合物能够转运或内化。因此,Ihog 在同种型粘附和配体结合中的看似不兼容的功能协同作用以协助 Hh 沿着纤毛发生进行运输和接收。