Department of Biochemistry, University of Oxford, Oxford, UK.
J Cell Biol. 2021 Aug 2;220(8). doi: 10.1083/jcb.202101022. Epub 2021 Jun 3.
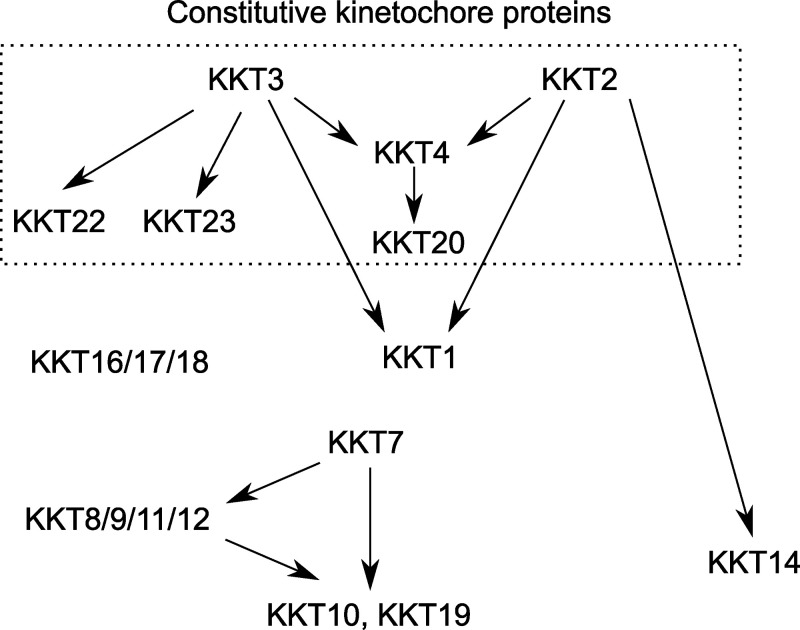
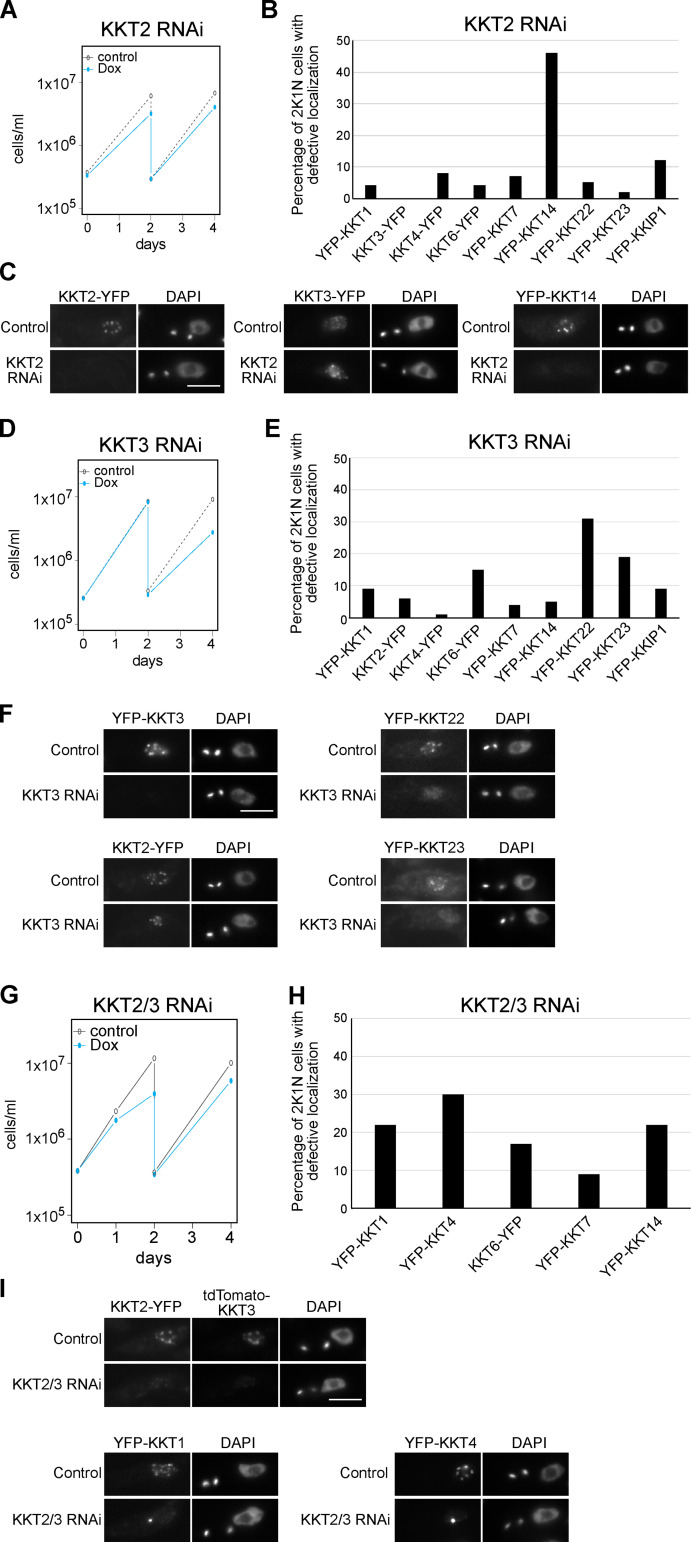
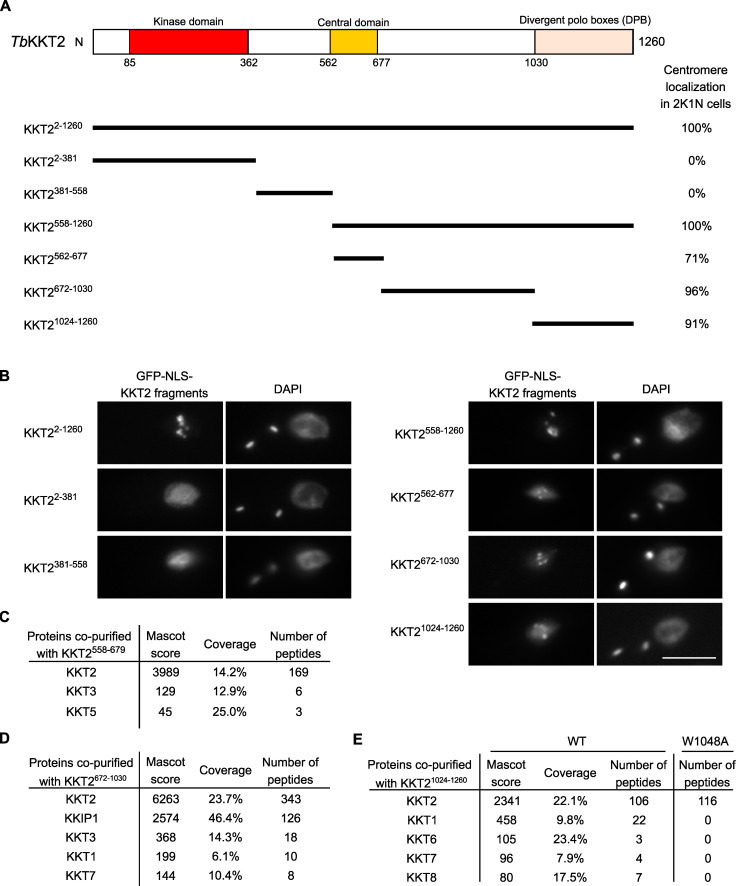
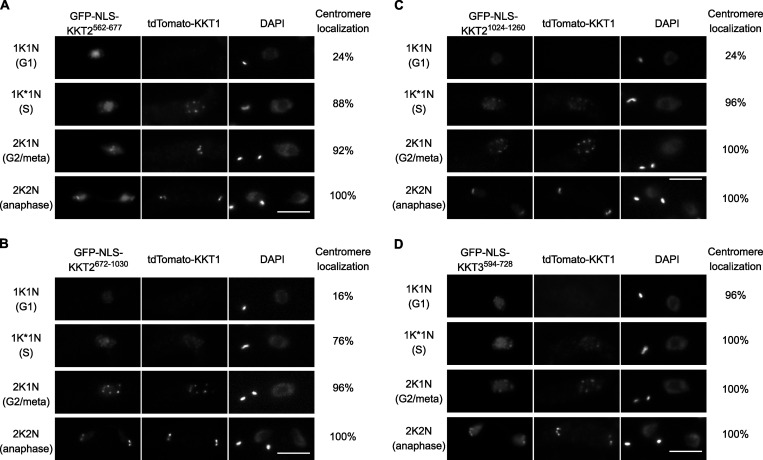
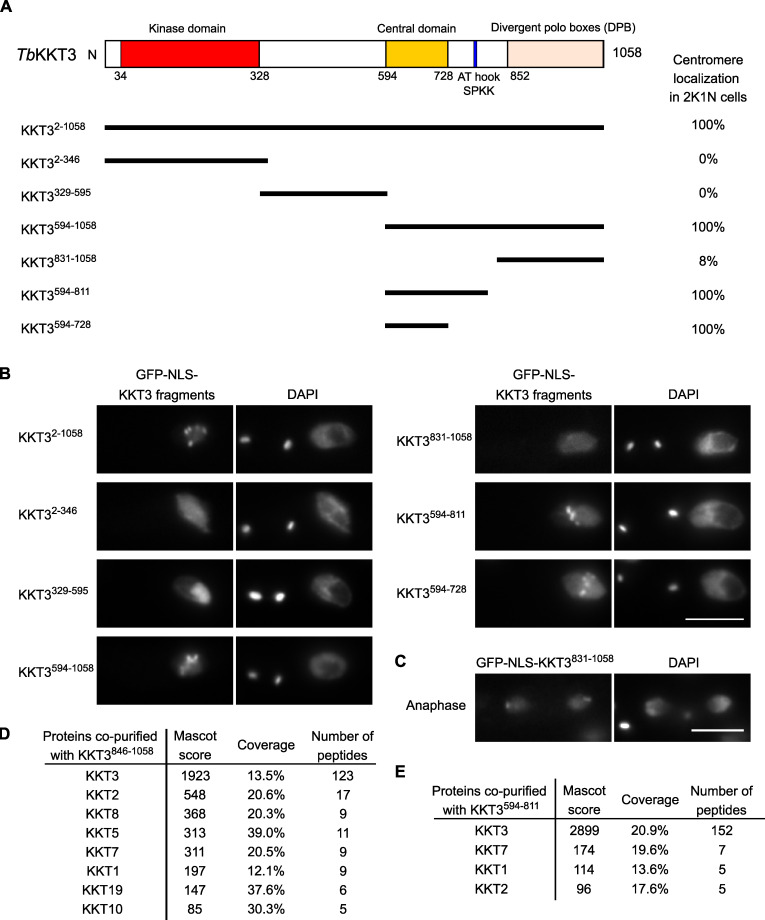
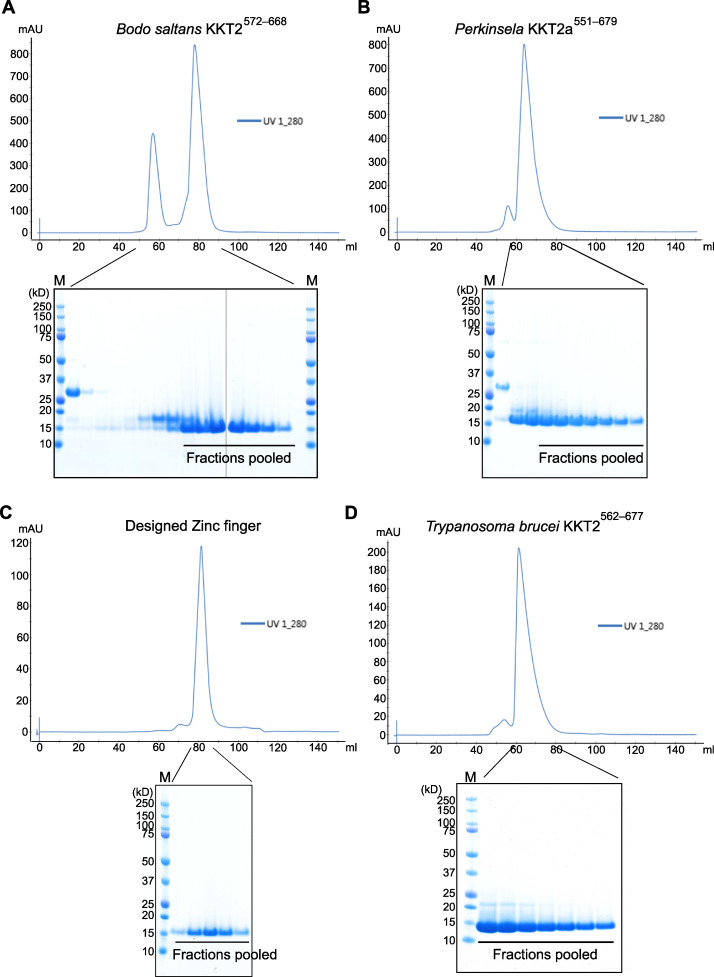
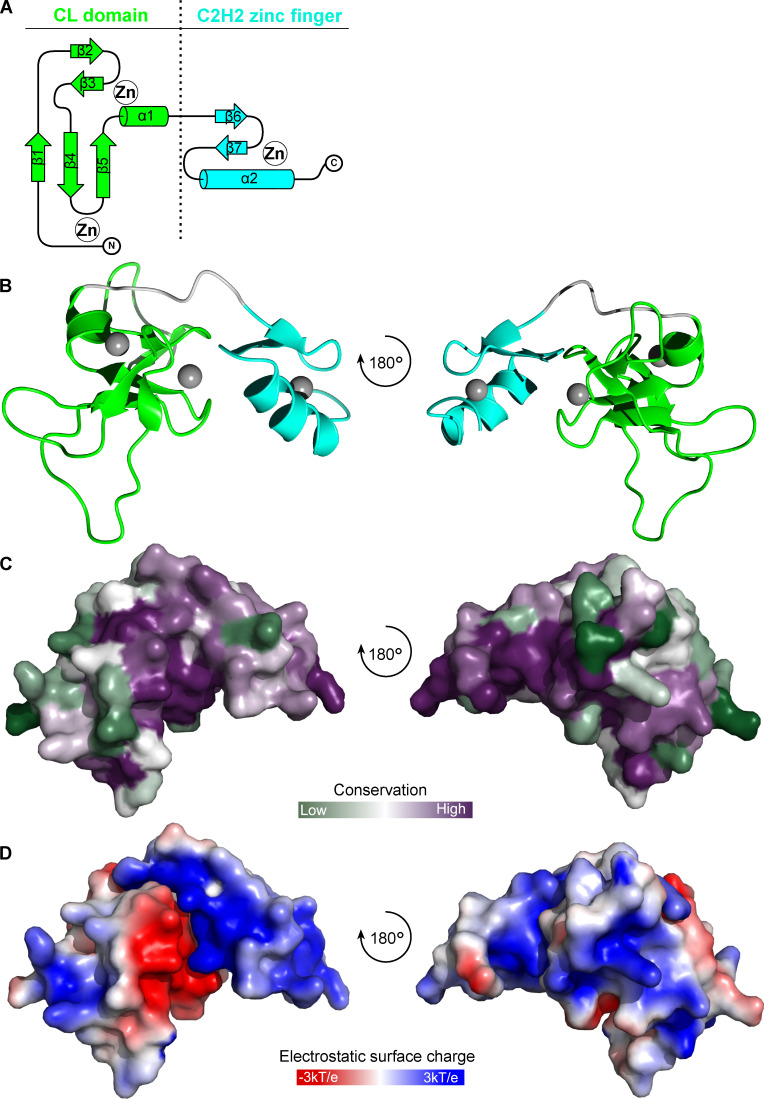
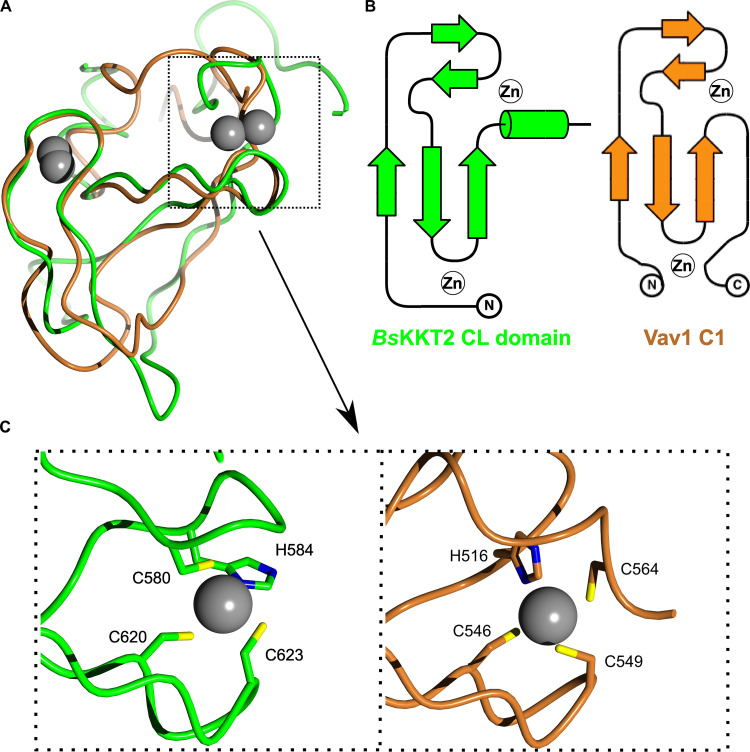
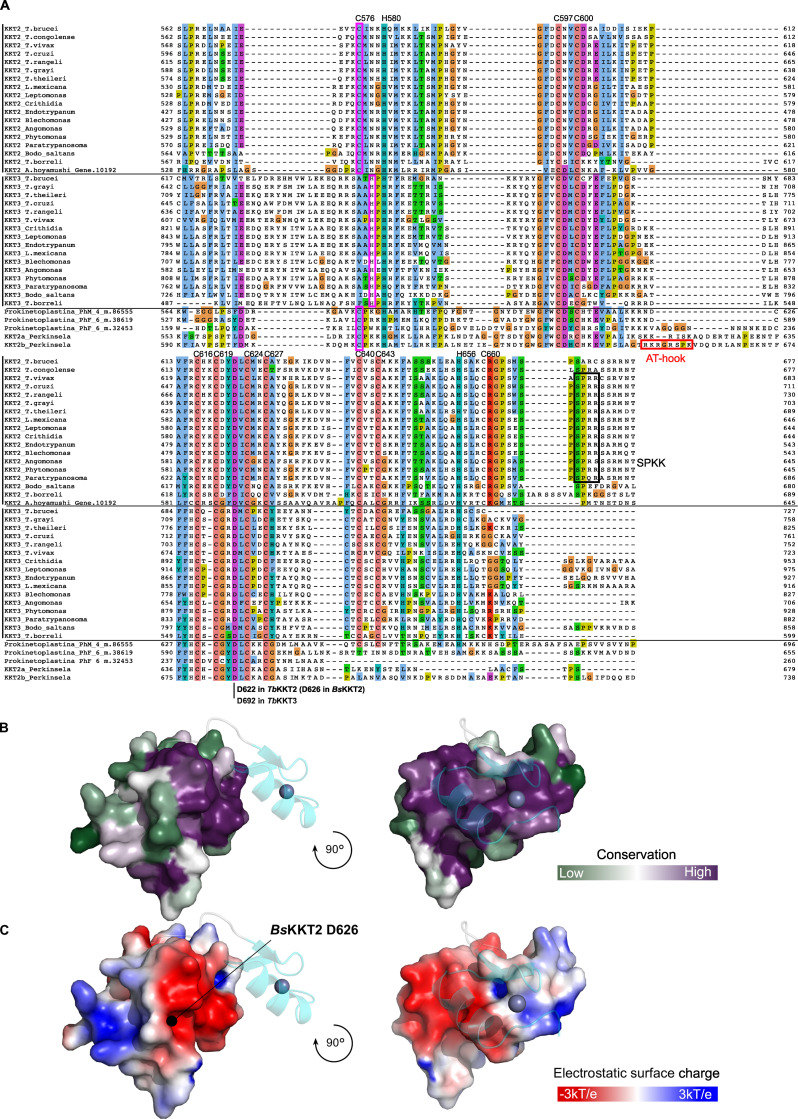
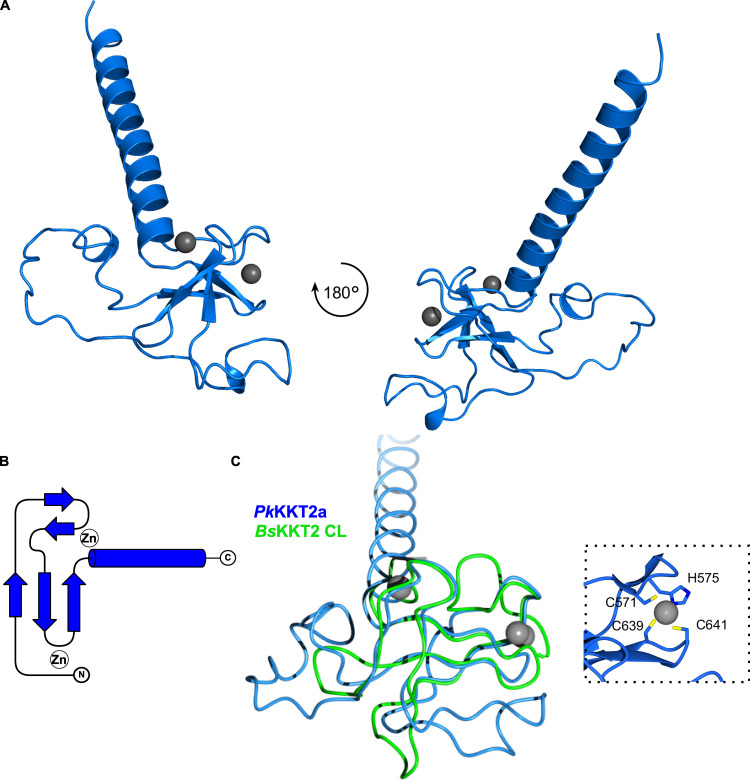
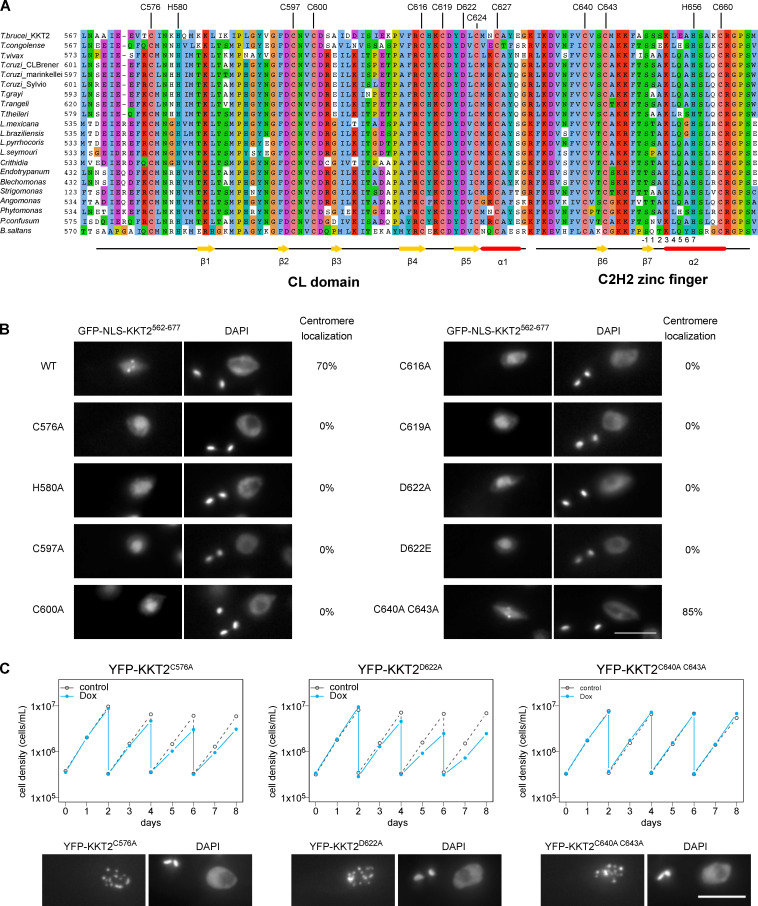
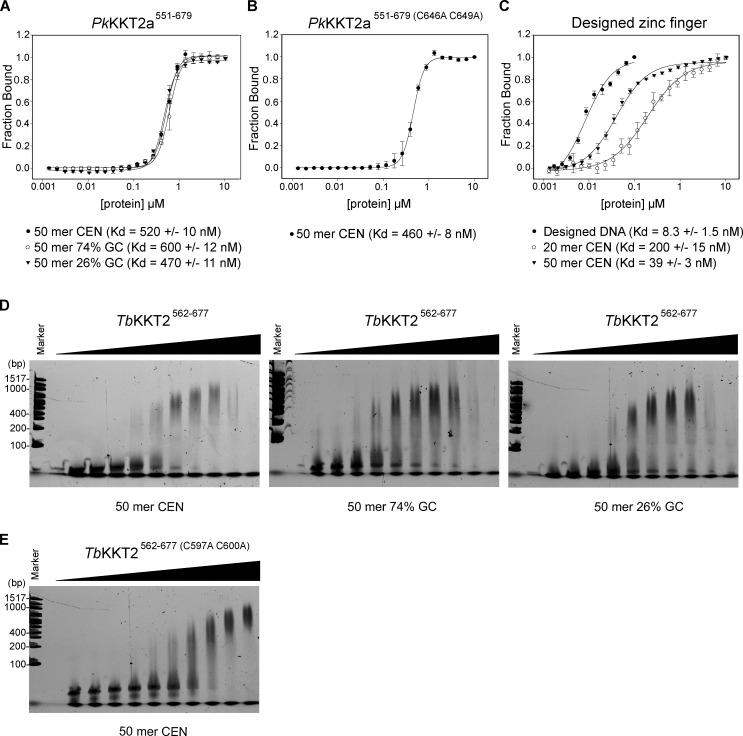
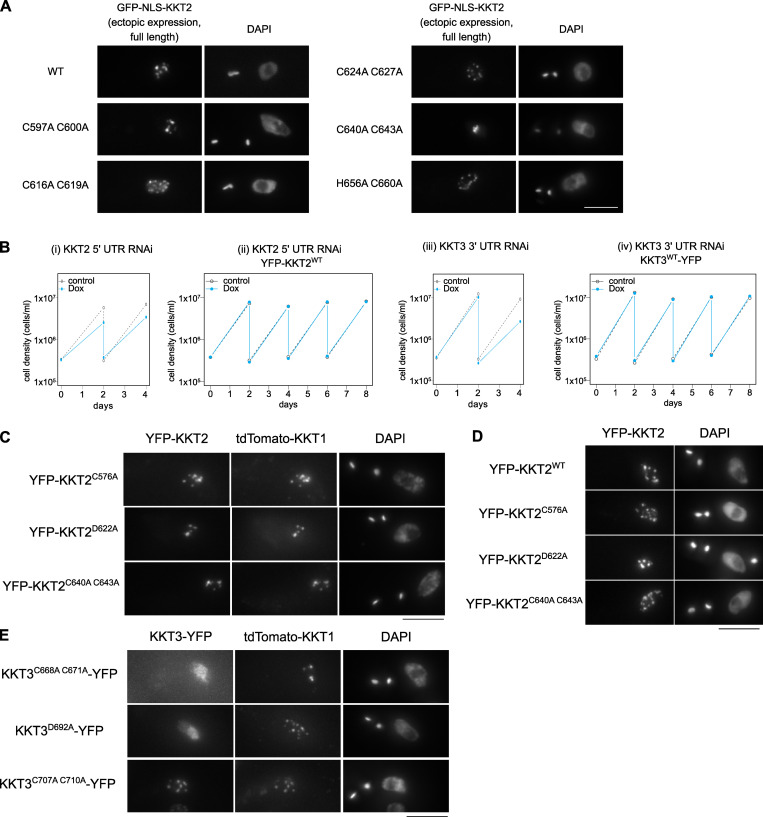
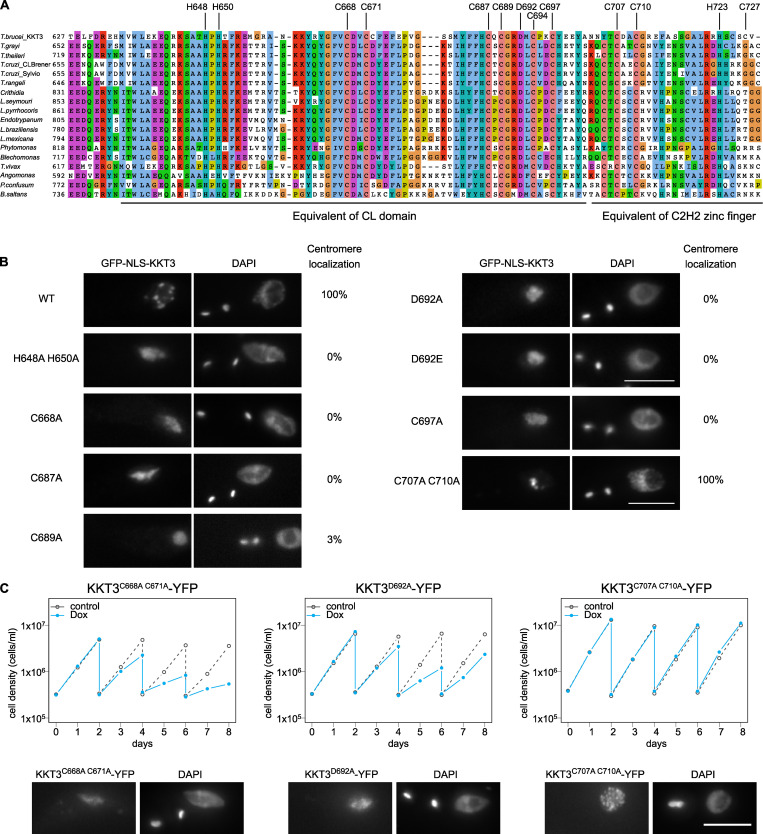
The kinetochore is the macromolecular protein complex that assembles onto centromeric DNA and binds spindle microtubules. Evolutionarily divergent kinetoplastids have an unconventional set of kinetochore proteins. It remains unknown how kinetochores assemble at centromeres in these organisms. Here, we characterize KKT2 and KKT3 in the kinetoplastid parasite Trypanosoma brucei. In addition to the N-terminal kinase domain and C-terminal divergent polo boxes, these proteins have a central domain of unknown function. We show that KKT2 and KKT3 are important for the localization of several kinetochore proteins and that their central domains are sufficient for centromere localization. Crystal structures of the KKT2 central domain from two divergent kinetoplastids reveal a unique zinc-binding domain (termed the CL domain for centromere localization), which promotes its kinetochore localization in T. brucei. Mutations in the equivalent domain in KKT3 abolish its kinetochore localization and function. Our work shows that the unique central domains play a critical role in mediating the centromere localization of KKT2 and KKT3.
着丝粒是组装在着丝粒 DNA 上并结合纺锤体微管的大分子蛋白复合物。进化上不同的动基体生物具有一组非常规的着丝粒蛋白。目前尚不清楚这些生物中的着丝粒如何在着丝粒上组装。在这里,我们对寄生虫锥虫中的动基体蛋白 KKT2 和 KKT3 进行了描述。除了 N 端激酶结构域和 C 端不同的 polo 盒结构域外,这些蛋白还具有一个功能未知的中心结构域。我们发现 KKT2 和 KKT3 对几种着丝粒蛋白的定位很重要,而且它们的中心结构域足以进行着丝粒定位。来自两种不同动基体生物的 KKT2 中心结构域的晶体结构揭示了一个独特的锌结合结构域(称为着丝粒定位的 CL 结构域),该结构域促进了其在 T. brucei 中的着丝粒定位。在 KKT3 中的等效结构域中的突变会使其丧失着丝粒定位和功能。我们的工作表明,独特的中心结构域在介导 KKT2 和 KKT3 的着丝粒定位中起着关键作用。