Department of Biochemistry and Molecular Biology, Johns Hopkins University Bloomberg School of Public Health, Baltimore, Maryland, USA.
Université Côte d'Azur, CNRS, INSERM, Institute of Biology Valrose (iBV), Parc Valrose, Nice, France.
J Biol Chem. 2021 Aug;297(2):100917. doi: 10.1016/j.jbc.2021.100917. Epub 2021 Jun 25.
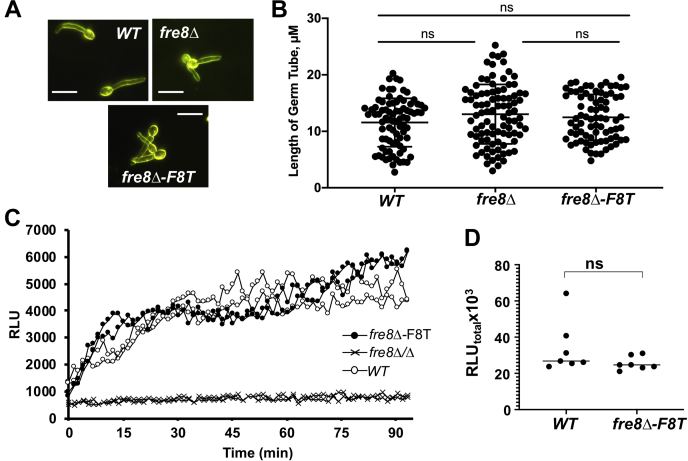
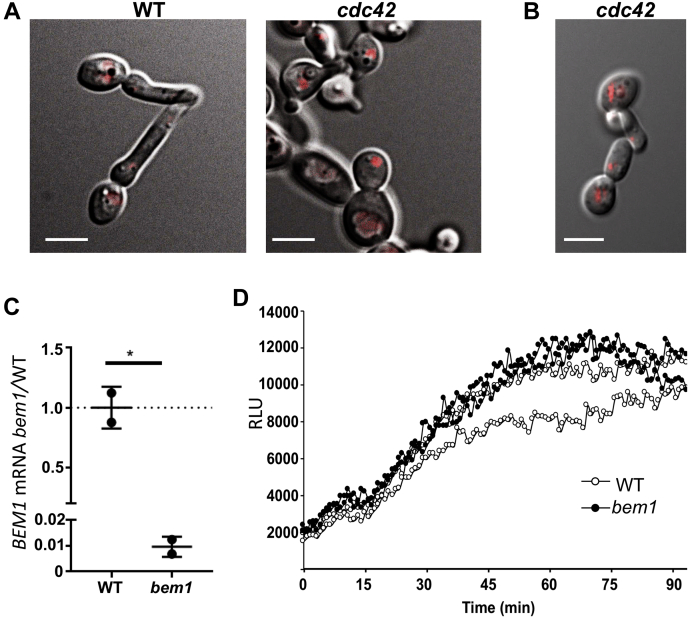
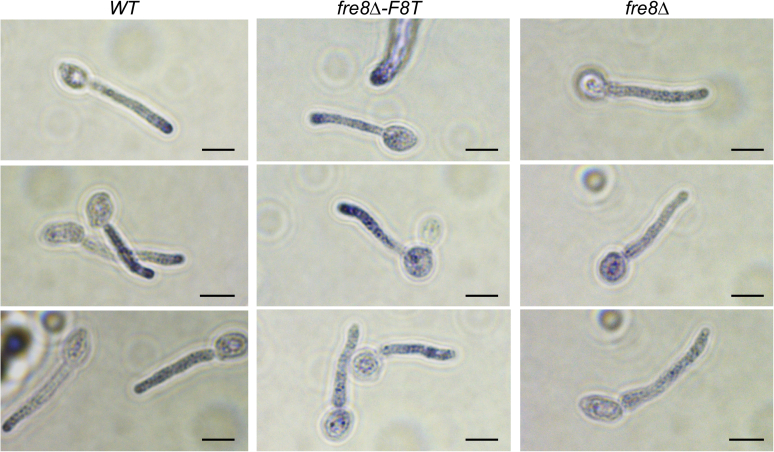
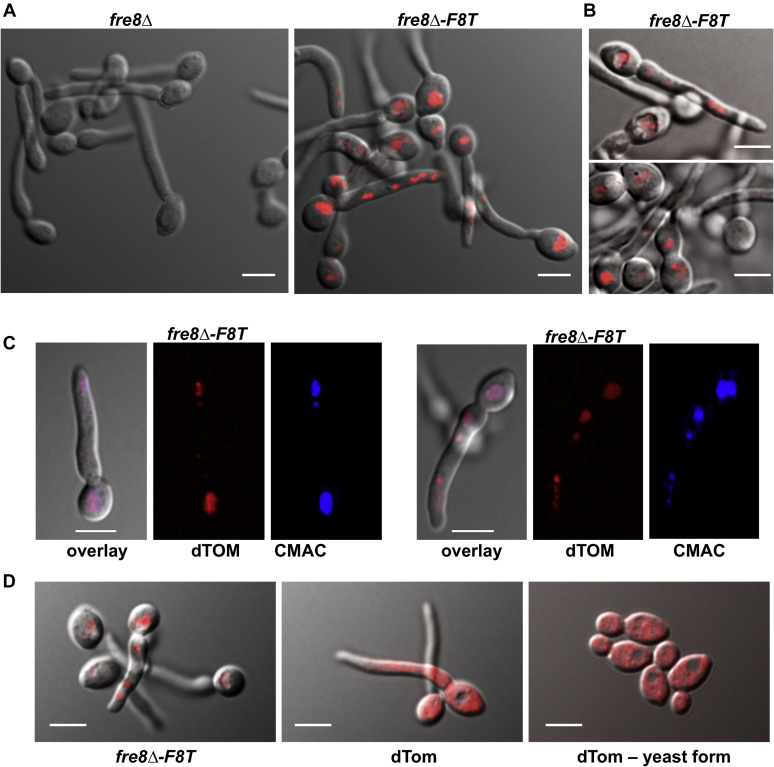
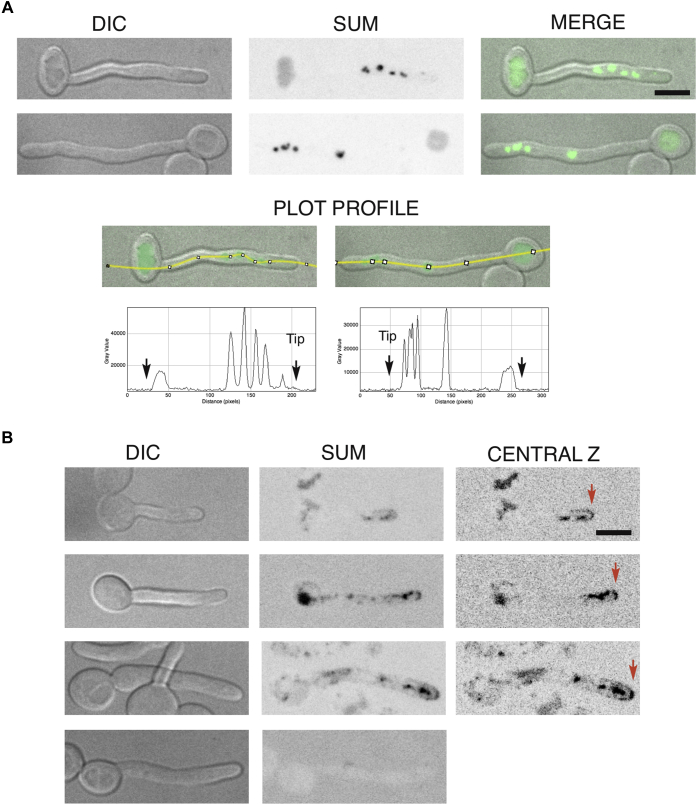
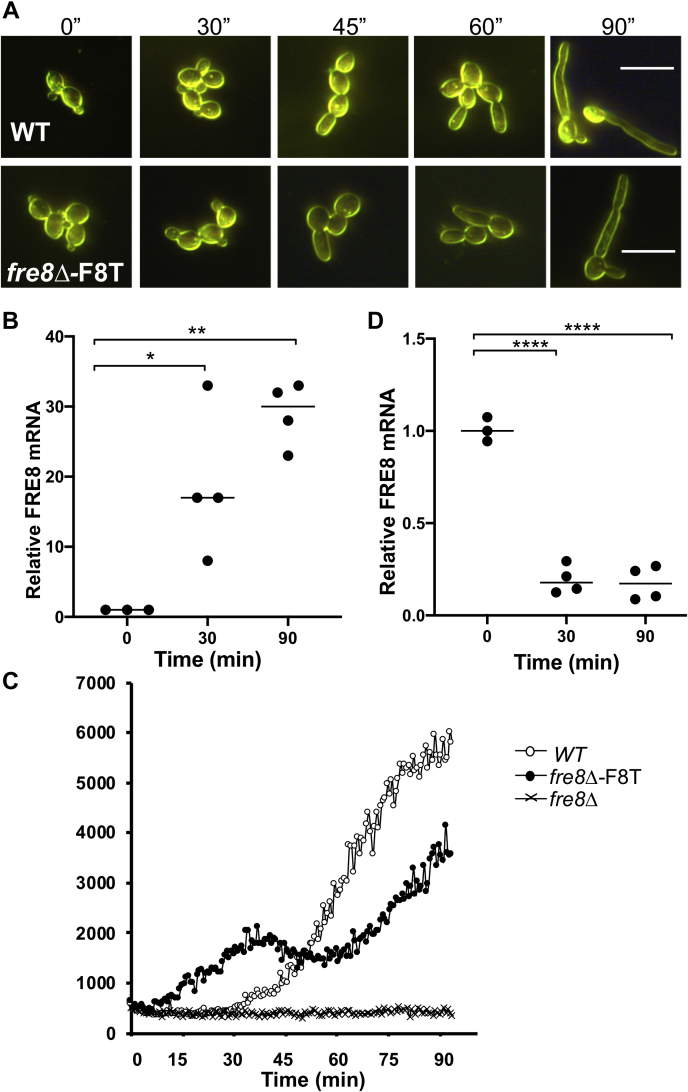
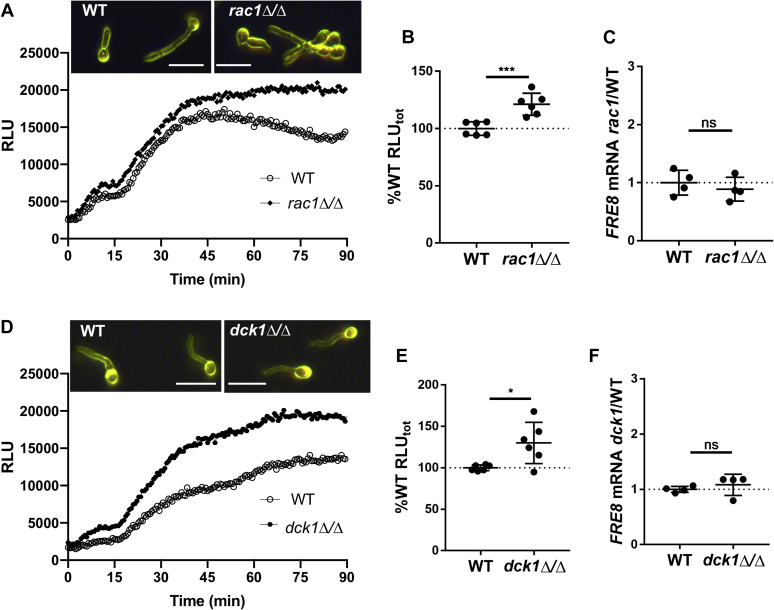
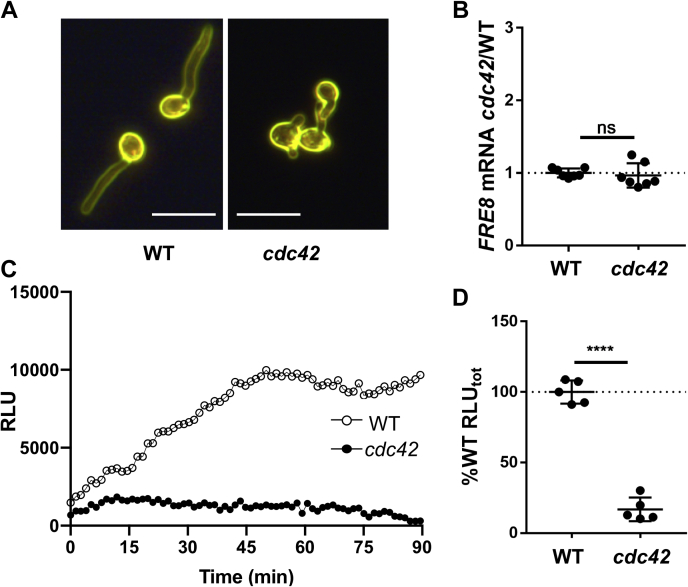
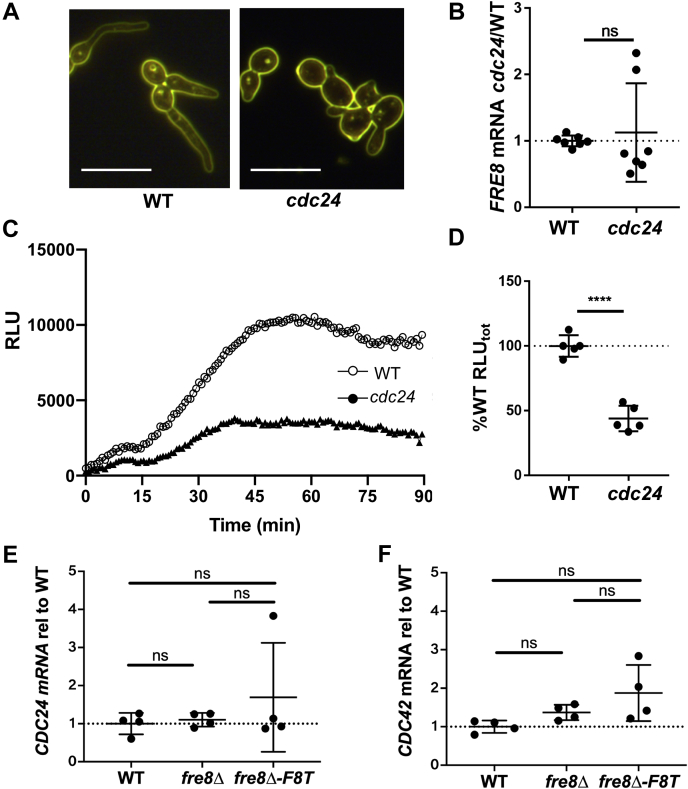
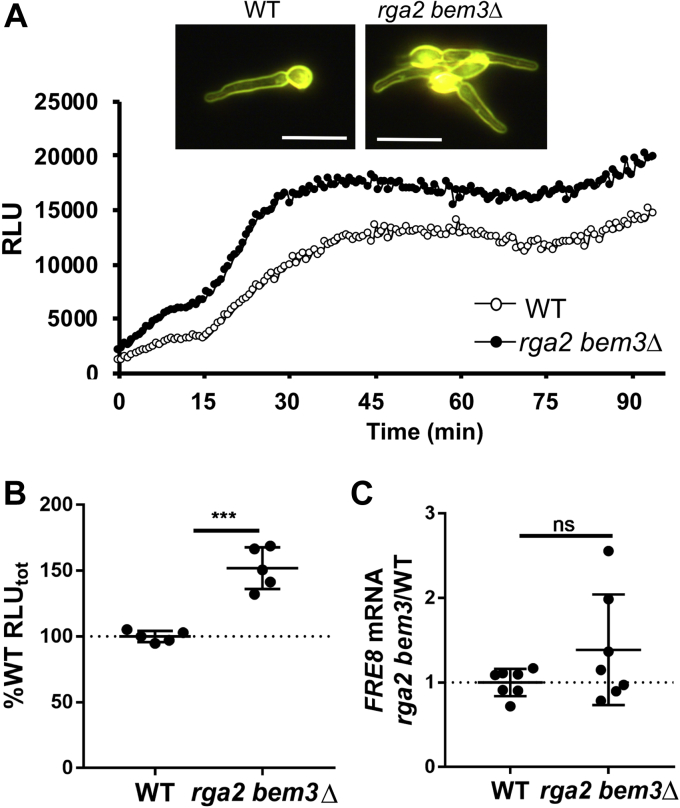
Across eukaryotes, Rho GTPases such as Rac and Cdc42 play important roles in establishing cell polarity, which is a key feature of cell growth. In mammals and filamentous fungi, Rac targets large protein complexes containing NADPH oxidases (NOX) that produce reactive oxygen species (ROS). In comparison, Rho GTPases of unicellular eukaryotes were believed to signal cell polarity without ROS, and it was unclear whether Rho GTPases were required for ROS production in these organisms. We document here the first example of Rho GTPase-mediated post-transcriptional control of ROS in a unicellular microbe. Specifically, Cdc42 is required for ROS production by the NOX Fre8 of the opportunistic fungal pathogen Candida albicans. During morphogenesis to a hyphal form, a filamentous growth state, C. albicans FRE8 mRNA is induced, which leads to a burst in ROS. Fre8-ROS is also induced during morphogenesis when FRE8 is driven by an ectopic promoter; hence, Fre8 ROS production is in addition controlled at the post-transcriptional level. Using fluorescently tagged Fre8, we observe that the majority of the protein is associated with the vacuolar system. Interestingly, much of Fre8 in the vacuolar system appears inactive, and Fre8-induced ROS is only produced at sites near the hyphal tip, where Cdc42 is also localized during morphogenesis. We observe that Cdc42 is necessary to activate Fre8-mediated ROS production during morphogenesis. Cdc42 regulation of Fre8 occurs without the large NOX protein complexes typical of higher eukaryotes and therefore represents a novel form of ROS control by Rho GTPases.
在真核生物中,Rho GTPases 如 Rac 和 Cdc42 在建立细胞极性方面发挥着重要作用,而细胞极性是细胞生长的关键特征。在哺乳动物和丝状真菌中,Rac 靶向包含 NADPH 氧化酶 (NOX) 的大型蛋白质复合物,这些复合物产生活性氧物种 (ROS)。相比之下,单细胞真核生物的 Rho GTPases 被认为在没有 ROS 的情况下发出细胞极性信号,并且不清楚 Rho GTPases 是否是这些生物体中 ROS 产生所必需的。我们在这里记录了 Rho GTPase 在单细胞微生物中调节 ROS 的转录后控制的第一个例子。具体来说,Cdc42 是机会性真菌病原体白色念珠菌的 NOX Fre8 产生 ROS 所必需的。在形态发生到菌丝形式,即丝状生长状态时,C. albicans FRE8 mRNA 被诱导,这导致 ROS 的爆发。当 FRE8 由异位启动子驱动时,也会在形态发生过程中诱导 Fre8-ROS;因此,Fre8 ROS 的产生还受到转录后水平的控制。使用荧光标记的 Fre8,我们观察到大多数蛋白质与液泡系统相关。有趣的是,液泡系统中的大部分 Fre8 似乎没有活性,并且只有在菌丝尖端附近的部位才会产生 Fre8 诱导的 ROS,在形态发生过程中,Cdc42 也定位于该部位。我们观察到 Cdc42 是在形态发生过程中激活 Fre8 介导的 ROS 产生所必需的。Cdc42 对 Fre8 的调节发生在没有高等真核生物中典型的大型 NOX 蛋白质复合物的情况下,因此代表了 Rho GTPases 对 ROS 控制的一种新形式。