Duhamel Aliénor, Benoit Julien, Day Michael, Rubidge Bruce, Fernandez Vincent
ENS de Lyon, CNRS, UMR 5276, LGL-TPE, Université Claude Bernard (Lyon I), Lyon, France.
Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa.
PeerJ. 2021 Aug 26;9:e11866. doi: 10.7717/peerj.11866. eCollection 2021.
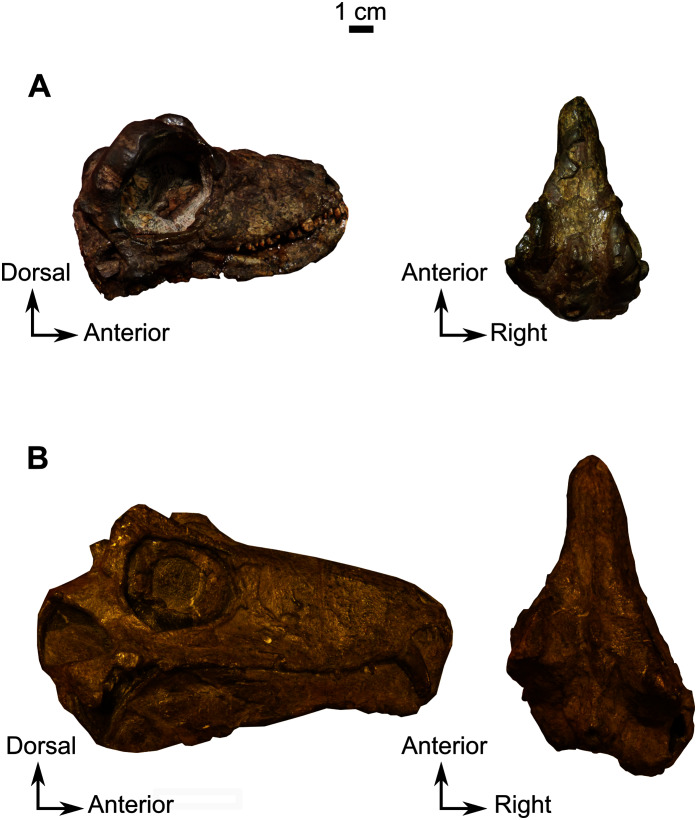
Biarmosuchia is a clade of basal therapsids that includes forms possessing plesiomorphic 'pelycosaurian' cranial characters as well as the highly derived Burnetiamorpha which are characterised by cranial pachyostosis and a variety of cranial bosses. Potential ontogenetic variation in these structures has been suggested based on growth series of other therapsids with pachyostosed crania, which complicates burnetiamorph taxonomic distinction and thus it is essential to better understand cranial ontogeny of the Burnetiamorpha. Here, three new juvenile biarmosuchian skulls from the late Permian of South Africa are described using X-ray micro computed tomography (CT). We found that juvenile biarmosuchians are distinguished from adults by their relatively large orbits, open cranial sutures, and incomplete ossification of the braincase and bony labyrinth. Also, they manifest multiple centres of ossification within the parietal and preparietal bones. CT examination reveals that the holotype of (BP/1/816), previously alleged to be a juvenile, shows no evidence of juvenility and is thus probably an adult. This suggests that the larger skull NMQR 1702, previously considered to be an adult , may represent a new taxon. This study provides, for the first time, a list of characters by which to recognise juvenile biarmosuchians.
二叠兽亚目是基干兽孔目的一个分支,包括具有原始“盘龙类”颅骨特征的类型以及高度特化的伯内蒂兽形类,其特征是颅骨骨质增生和各种颅骨隆起。基于其他具有骨质增生颅骨的兽孔类动物的生长序列,有人提出了这些结构中潜在的个体发育变异,这使得伯内蒂兽形类的分类区分变得复杂,因此,更好地了解伯内蒂兽形类的颅骨个体发育至关重要。在此,利用X射线显微计算机断层扫描(CT)描述了来自南非晚二叠世的三个新的幼年二叠兽亚目头骨。我们发现,幼年二叠兽亚目与成年个体的区别在于其相对较大的眼眶、开放的颅骨缝线以及脑壳和骨迷路的不完全骨化。此外,它们在顶骨和前顶骨内表现出多个骨化中心。CT检查显示,以前被认为是幼年个体的(BP/1/816)的正模标本没有幼年特征的证据,因此可能是成年个体。这表明,以前被认为是成年个体的较大的头骨NMQR 1702可能代表一个新分类单元。这项研究首次提供了识别幼年二叠兽亚目的一系列特征。