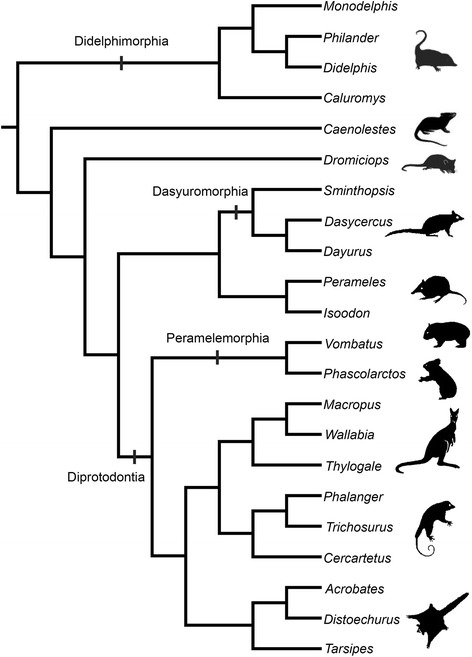
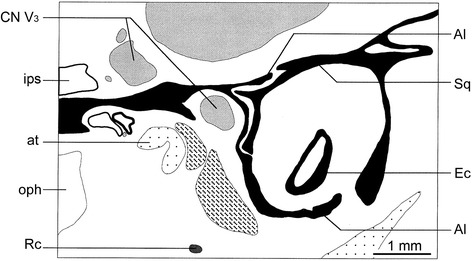
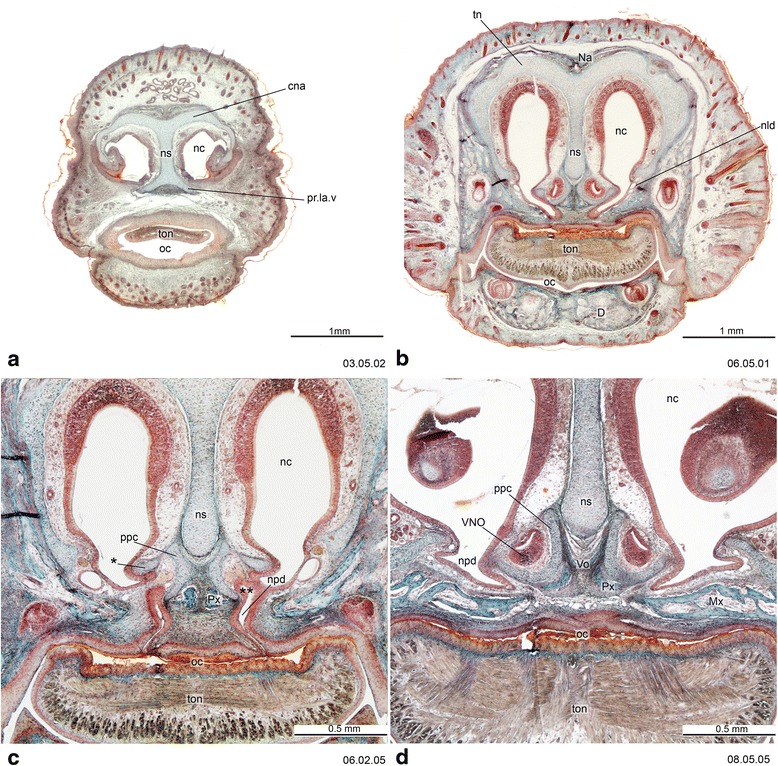
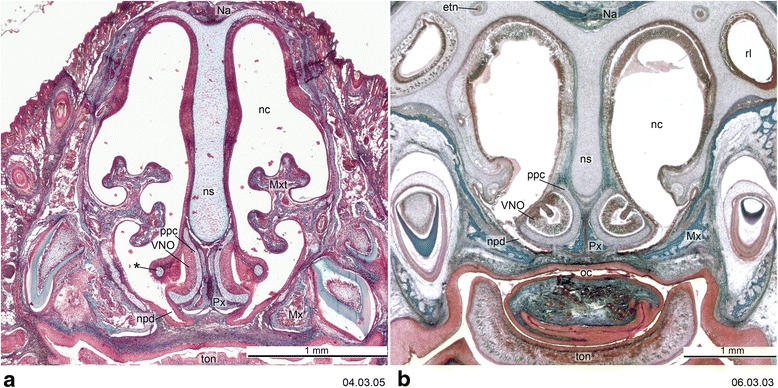
Sánchez-Villagra Marcelo R, Forasiepi Analía M
Paläontologisches Institut und Museum der Universität Zürich, Karl Schmid Strasse 4, Zürich, 8006 Switzerland.
IANIGLA, CCT-CONICET Mendoza, Av. Ruiz Leal s/n°, Mendoza, 5500 Argentina.
Zoological Lett. 2017 Feb 12;3:1. doi: 10.1186/s40851-017-0062-y. eCollection 2017.
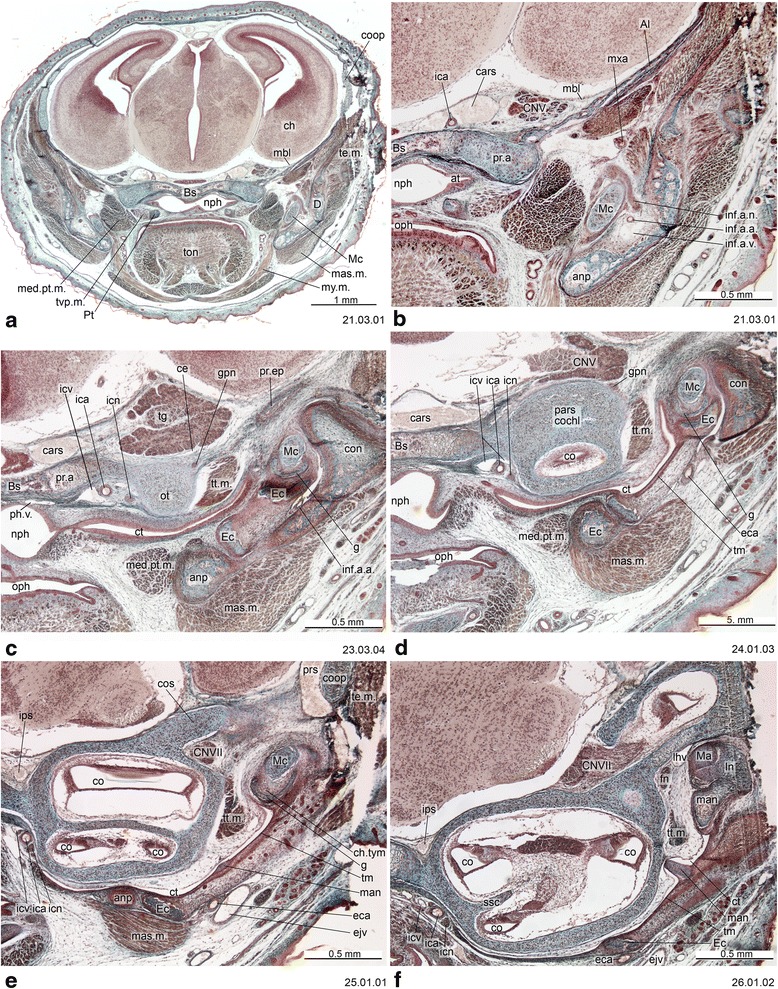
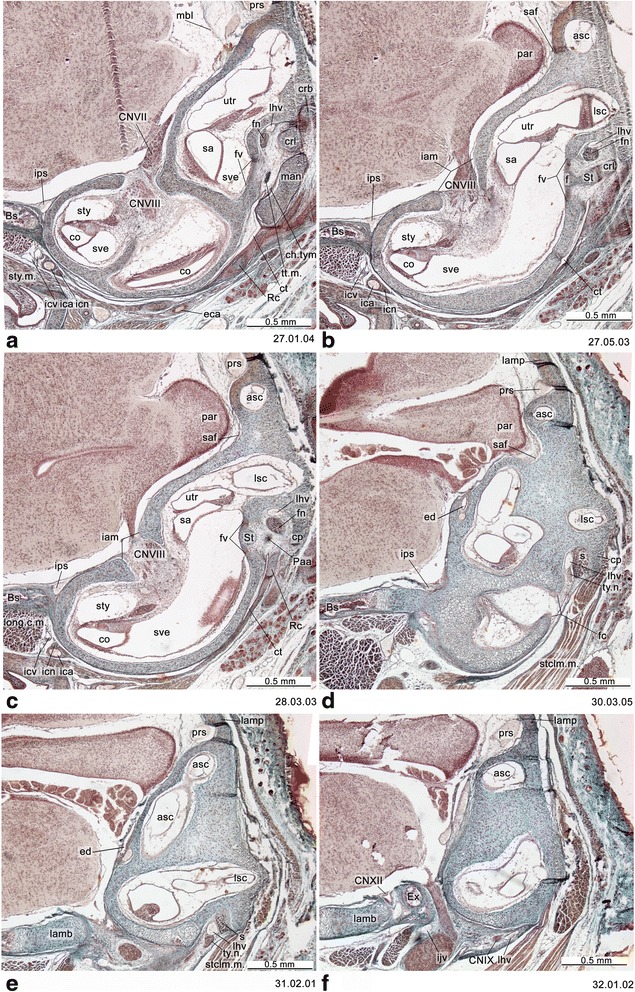
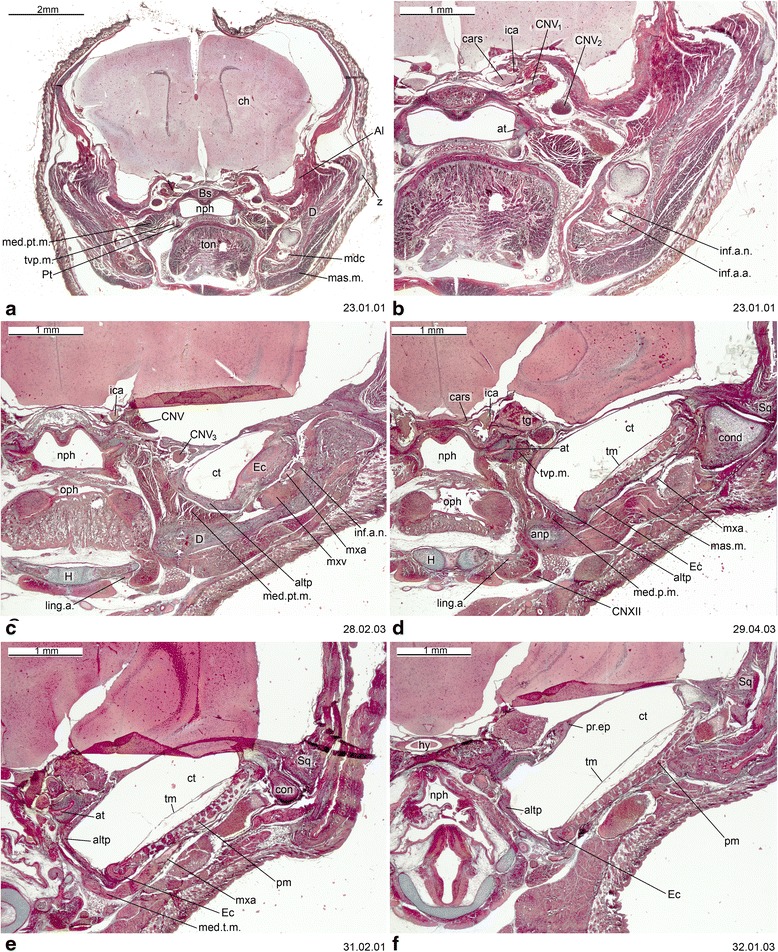
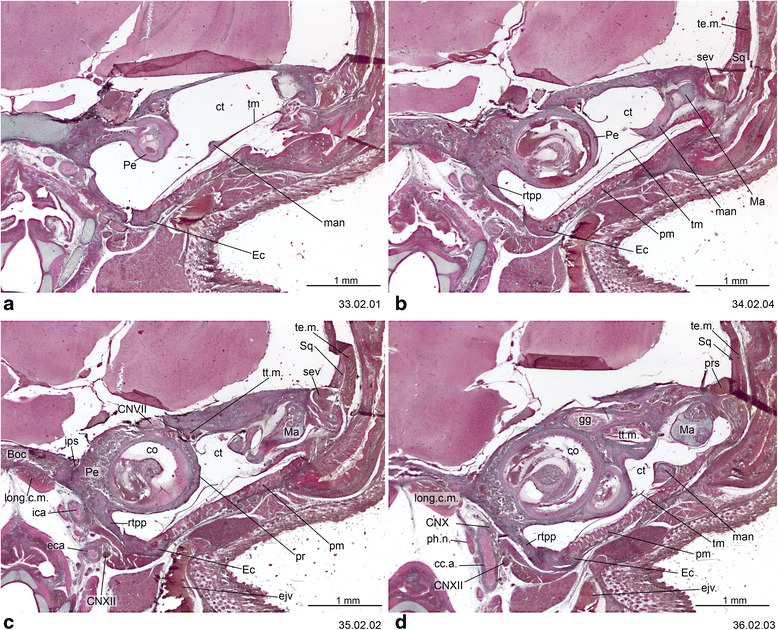
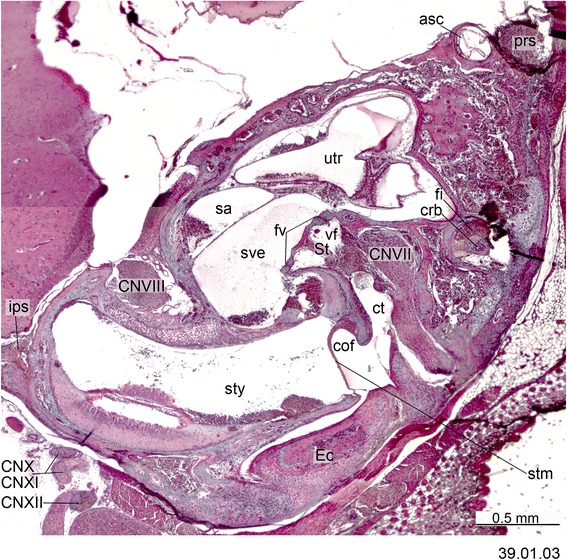
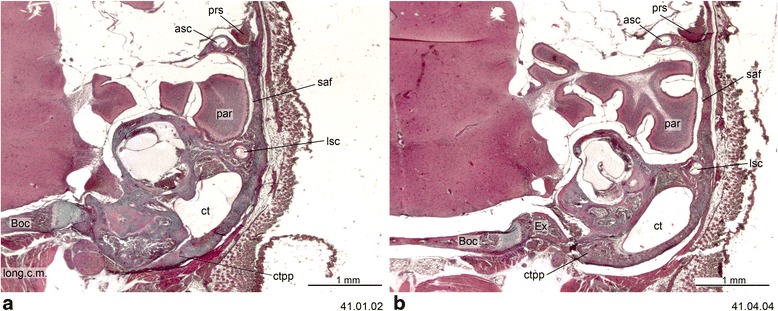
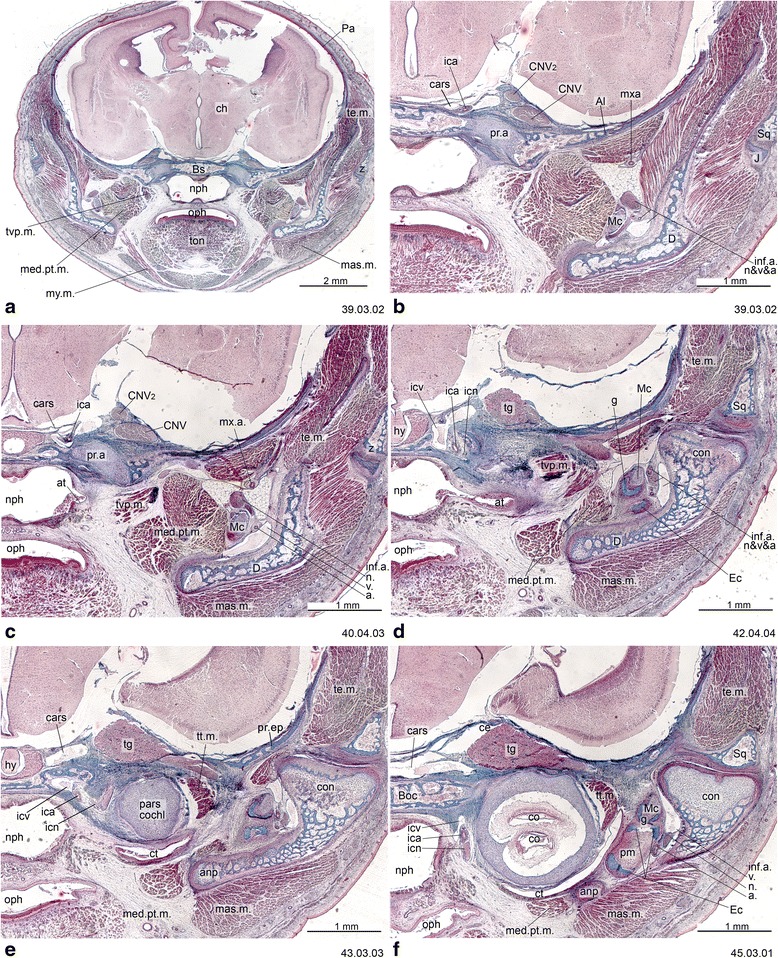
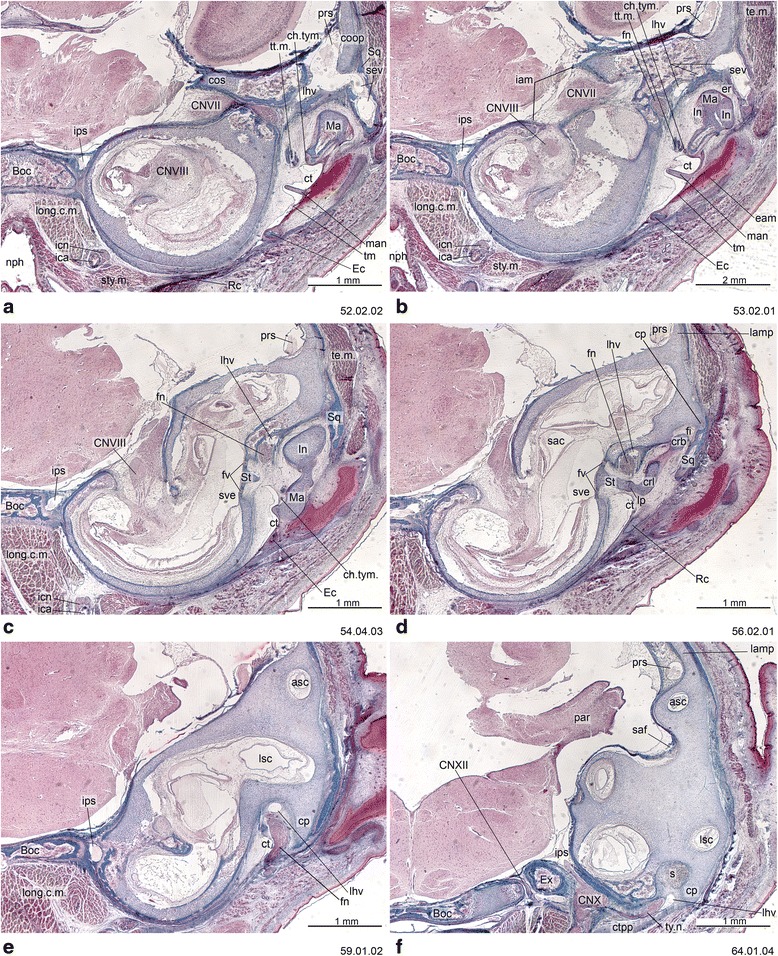
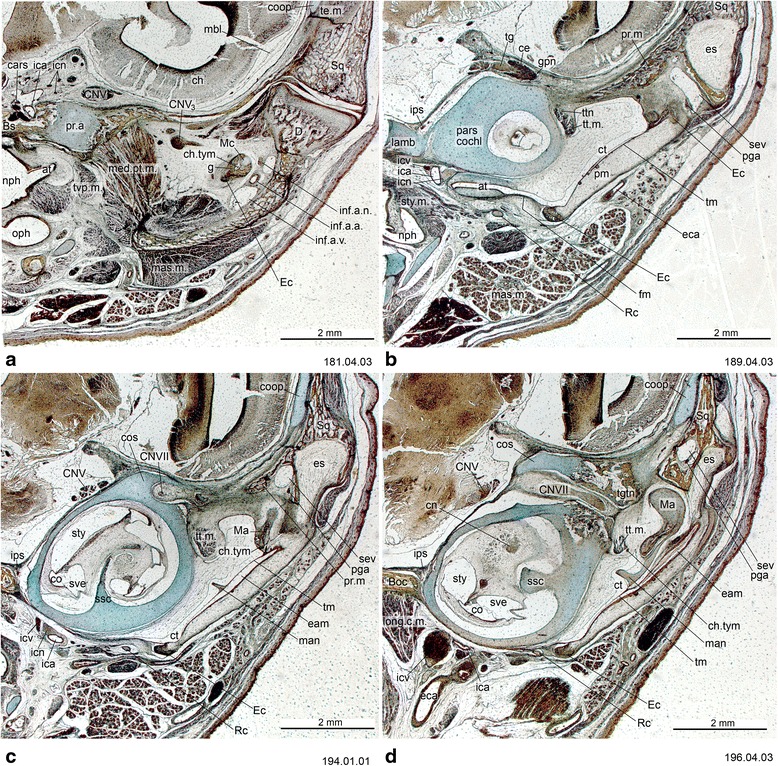
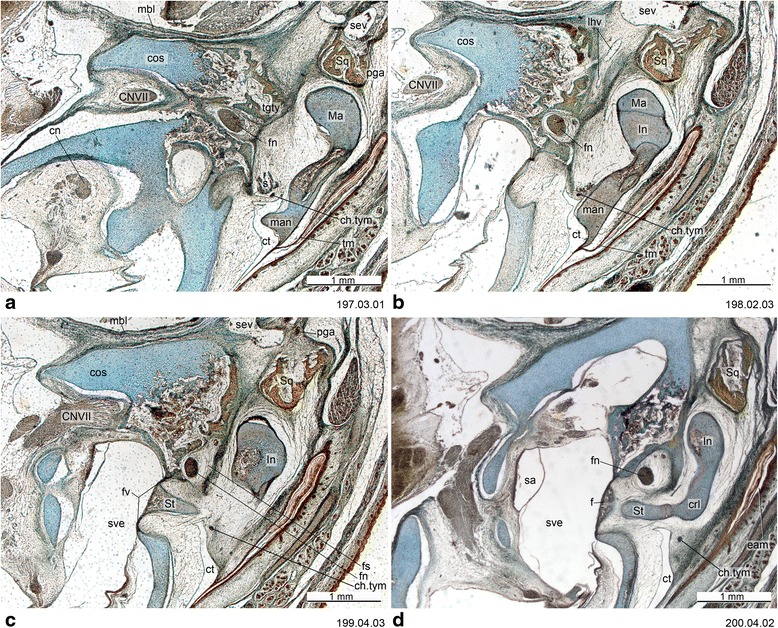
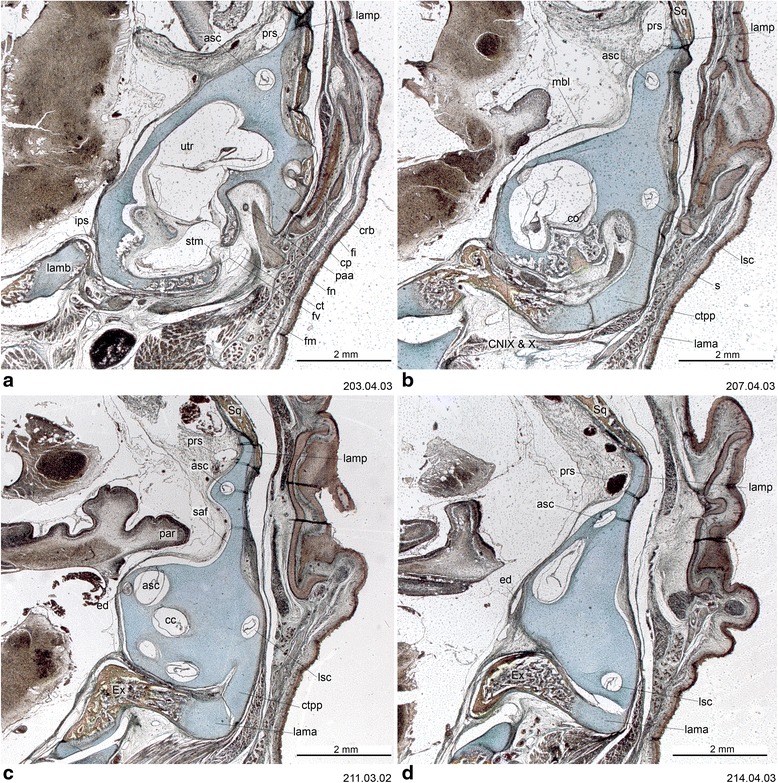
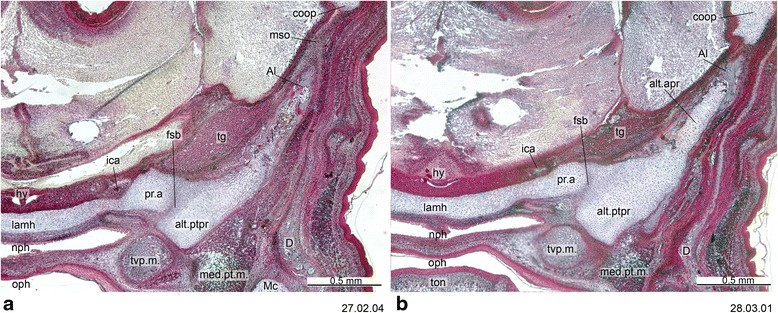
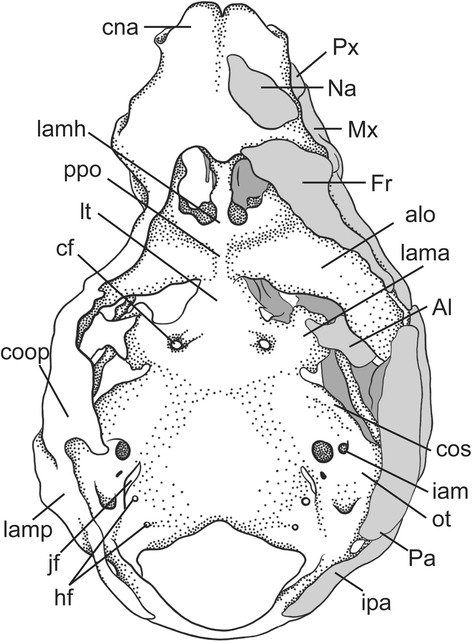
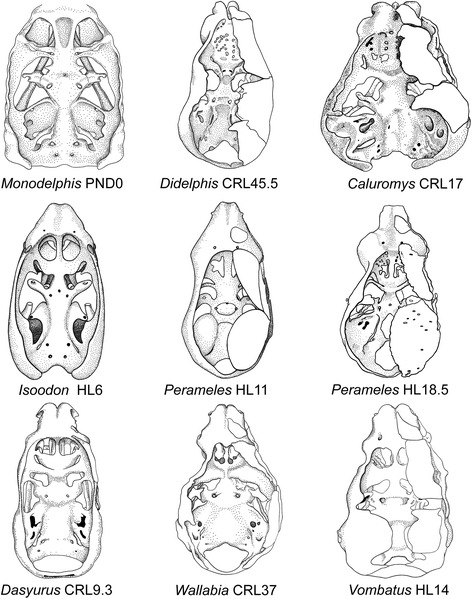
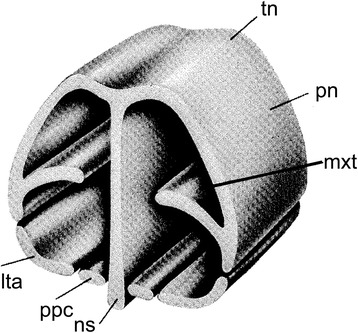
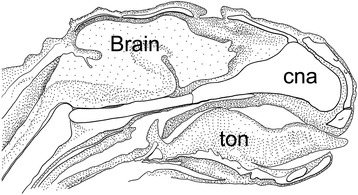
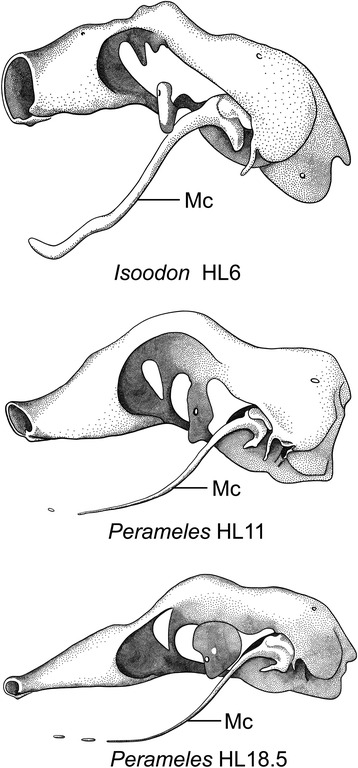
An overview of the literature on the chondrocranium of marsupial mammals reveals a relative conservatism in shape and structures. We document the histological cranial anatomy of individuals representing , , sp. and . The marsupial chondrocranium is generally characterized by the great breadth of the lamina basalis, absence of pila metoptica and large otic capsules. Its most anterior portion (cupula nasi anterior) is robust, and anterior to it there are well-developed tactile sensory structures, functionally important in the neonate. Investigations of ossification centers at and around the nasal septum are needed to trace the presence of certain bones (e.g., mesethmoid, parasphenoid) across marsupial taxa. In many adult marsupials, the tympanic floor is formed by at least three bones: alisphenoid (alisphenoid tympanic process), ectotympanic and petrosal (rostral and caudal tympanic processes); the squamosal also contributes in some diprotodontians. The presence of an entotympanic in marsupials has not been convincingly demonstrated. The tubal element surrounding the auditory tube in most marsupials is fibrous connective tissue rather than cartilage; the latter is the case in most placentals recorded to date. However, we detected fibrocartilage in a late juvenile of , and a similar tissue has been reported for . Contradictory reports on the presence of the tegmen tympani can be found in the literature. We describe a small tegmen tympani in . Several heterochronic shifts in the timing of development of the chondocranium and associated structures (e.g., nerves, muscles) and in the ossification sequence have been interpreted as largely being influenced by functional requirements related to the altriciality of the newborn marsupial during early postnatal life. Comparative studies of chondocranial development of mammals can benefit from a solid phylogenetic framework, research on non-classical model organisms, and integration with imaging and sectional data derived from computer-tomography.
有袋类哺乳动物软骨颅的文献综述显示,其形状和结构相对保守。我们记录了代表 、 、 属和 属的个体的组织学颅部解剖结构。有袋类软骨颅的一般特征是基底板宽阔、无额顶柱且耳囊大。其最前部(前鼻穹窿)粗壮,在其前方有发育良好的触觉感觉结构,这在新生儿中具有重要功能。需要对鼻中隔及其周围的骨化中心进行研究,以追踪某些骨骼(如中筛骨、副蝶骨)在有袋类分类群中的存在情况。在许多成年有袋类动物中,鼓膜底至少由三块骨头形成:翼蝶骨(翼蝶骨鼓膜突)、鼓骨和岩骨(鼓膜前后突);在一些双门齿目动物中,鳞骨也有贡献。有袋类动物中鼓室内骨的存在尚未得到令人信服的证实。大多数有袋类动物中围绕咽鼓管的管道部分是纤维结缔组织而非软骨;而迄今为止记录的大多数胎盘类动物情况则相反。然而,我们在一只 的晚期幼体中检测到了纤维软骨,并且已有报道在 中也存在类似组织。关于鼓室盖的存在,文献中有相互矛盾的报道。我们描述了 在 中有一个小的鼓室盖。软骨颅及其相关结构(如神经、肌肉)发育时间和骨化序列的几个异时性变化在很大程度上被解释为受到与新生有袋类动物出生后早期的未成熟状态相关的功能需求的影响。哺乳动物软骨颅发育的比较研究可以受益于坚实的系统发育框架、对非经典模式生物的研究以及与计算机断层扫描获得的成像和断层数据的整合。