Department of Chemistry, University of Washington, Seattle, United States.
Laboratory of Biochemistry and Molecular Biology, The Rockefeller University, New York City, United States.
Elife. 2021 Nov 8;10:e67952. doi: 10.7554/eLife.67952.
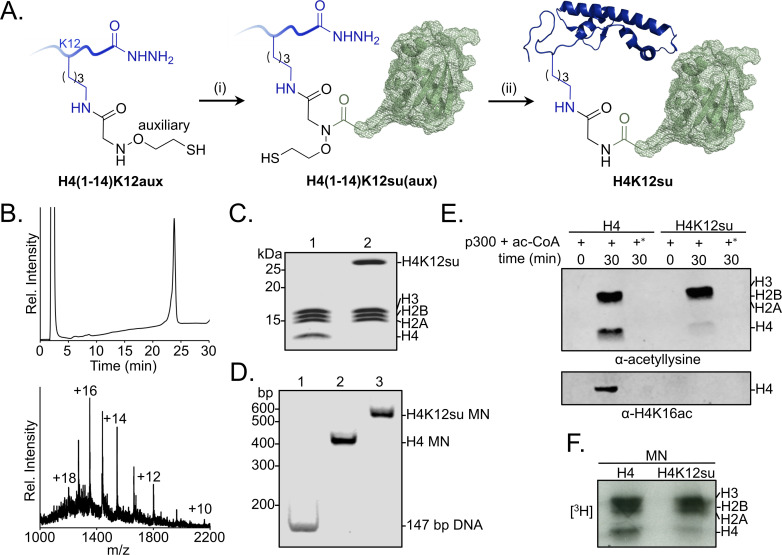
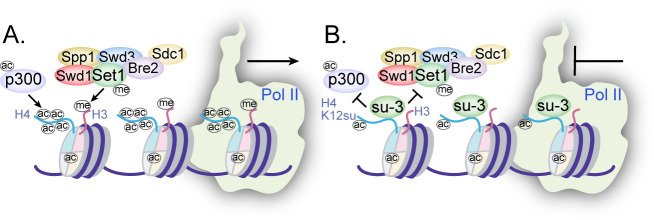
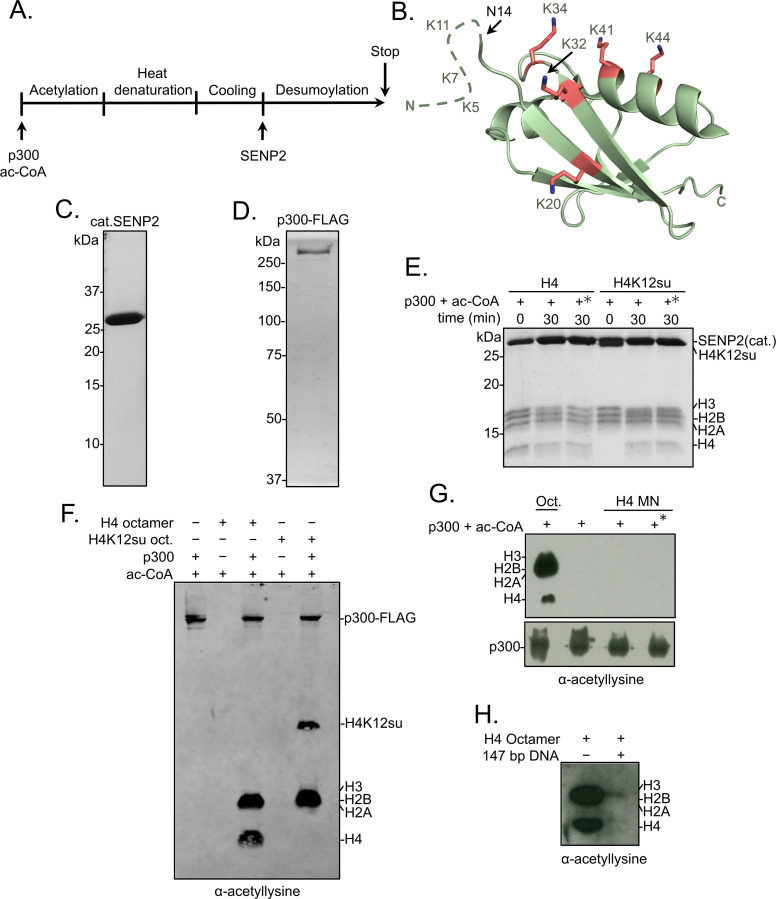
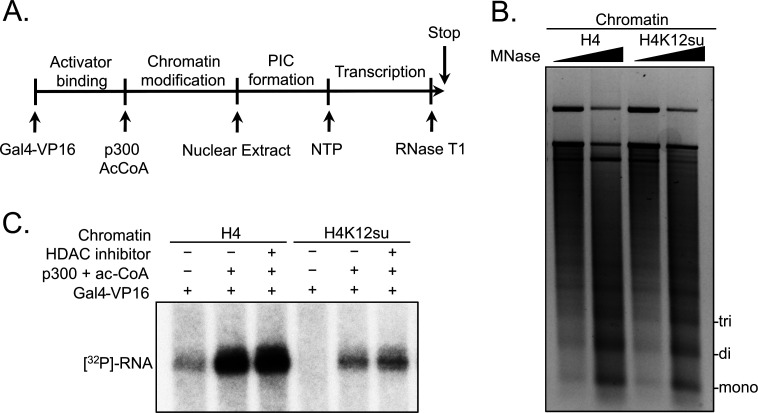
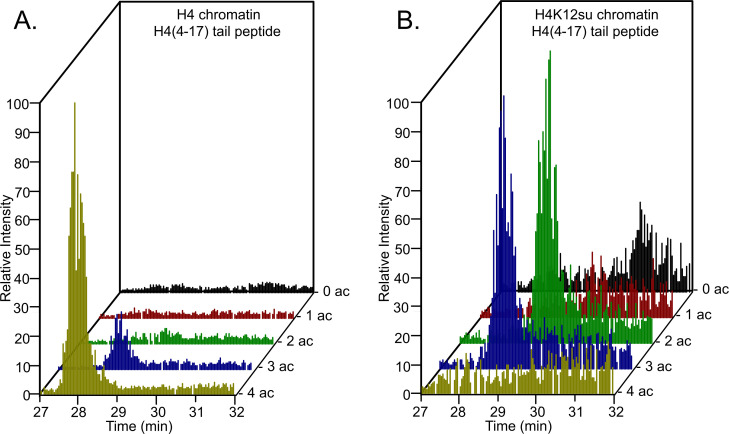
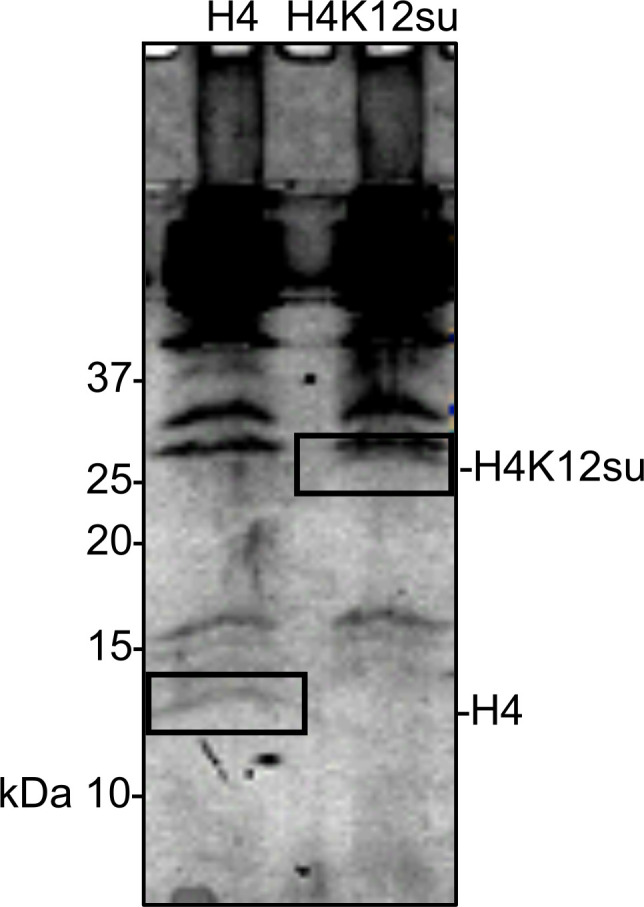
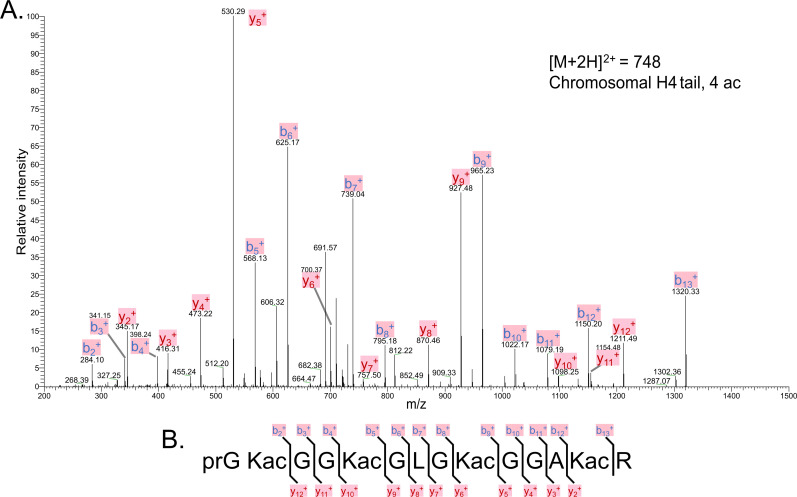
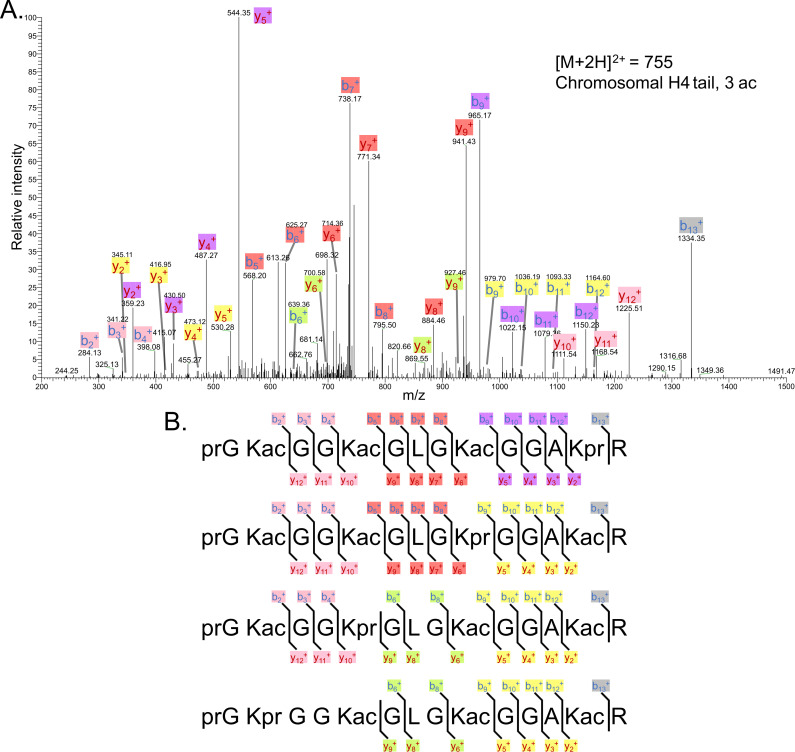
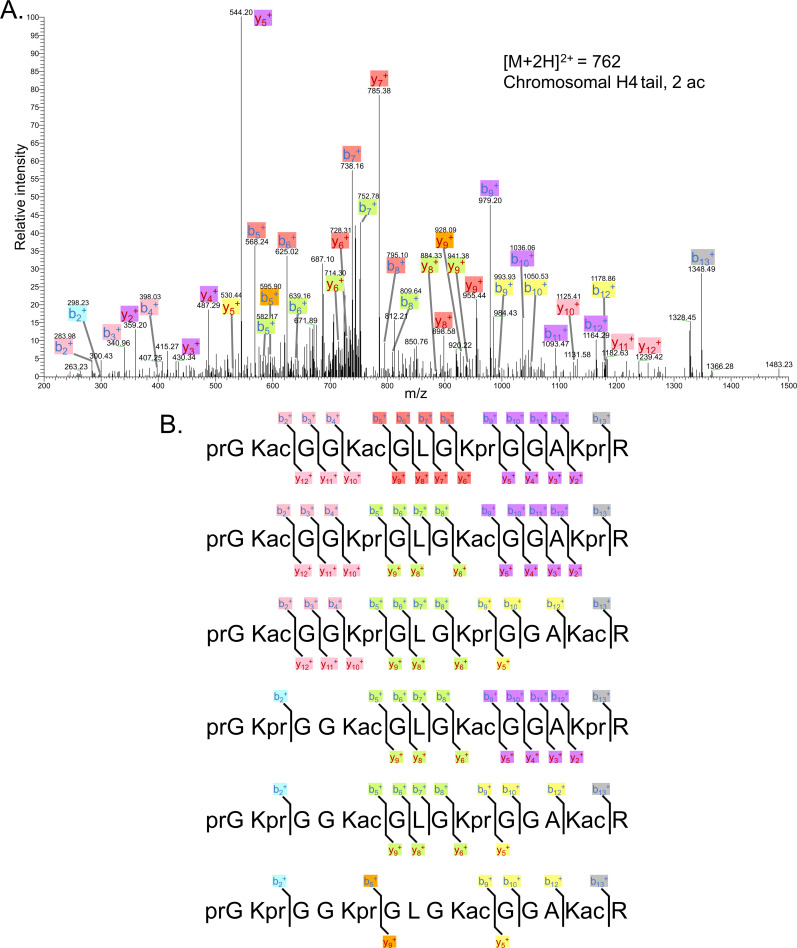
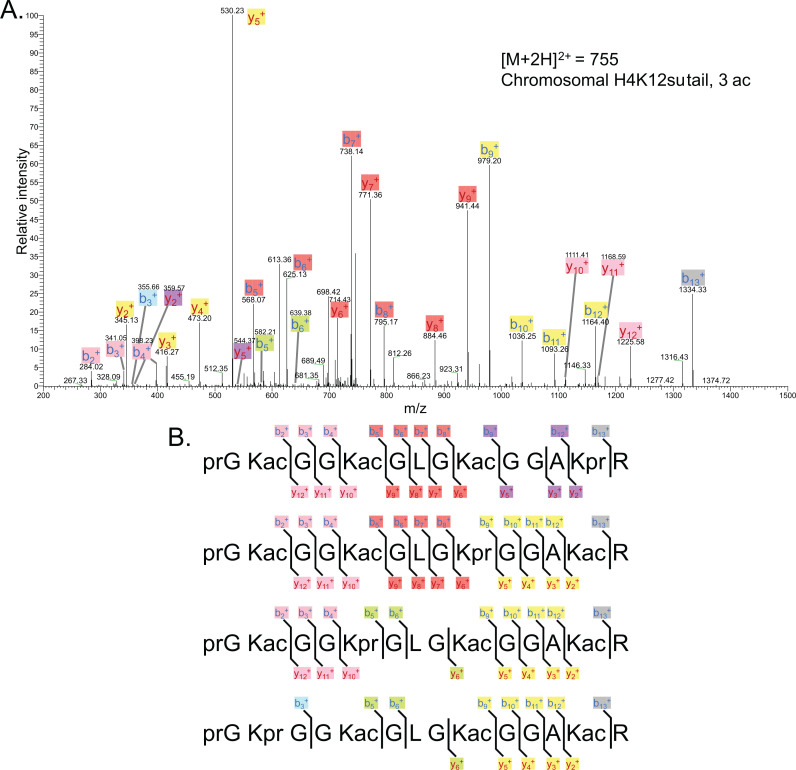
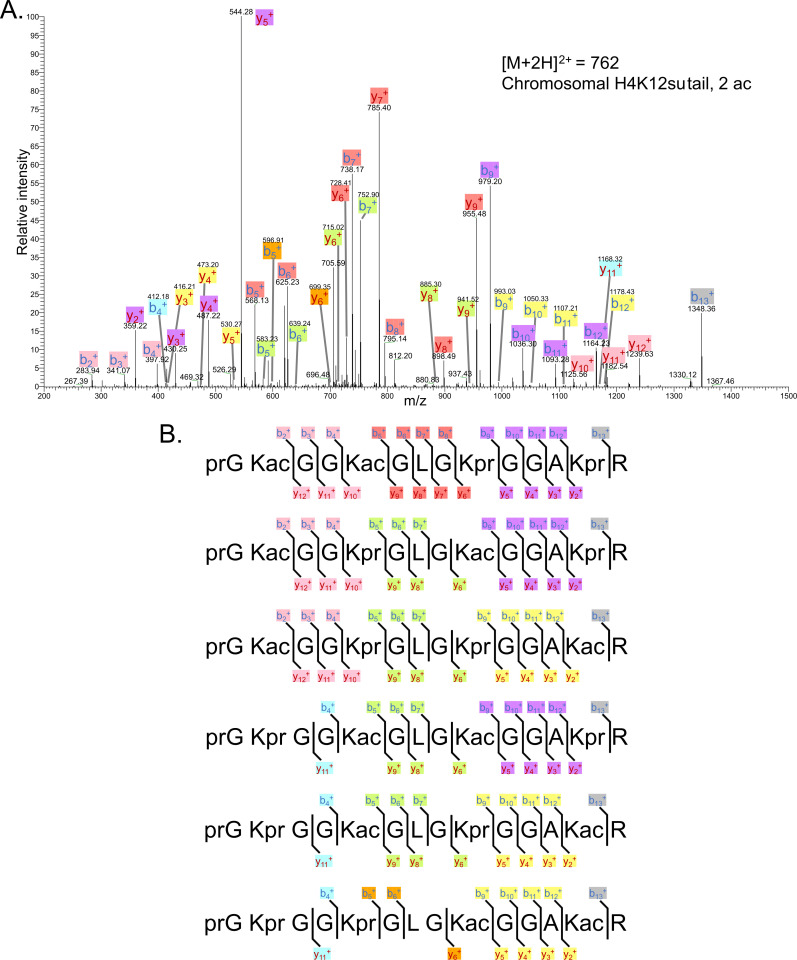
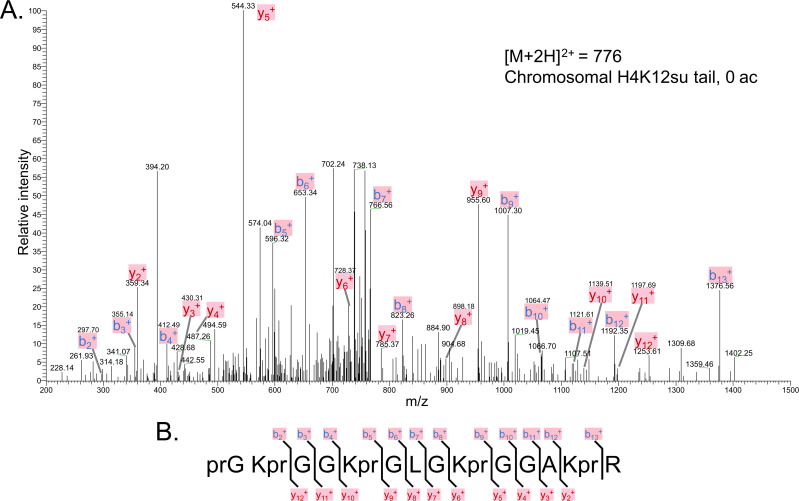
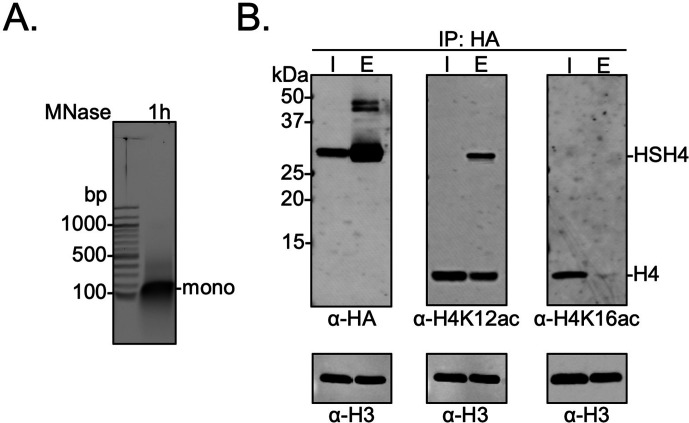
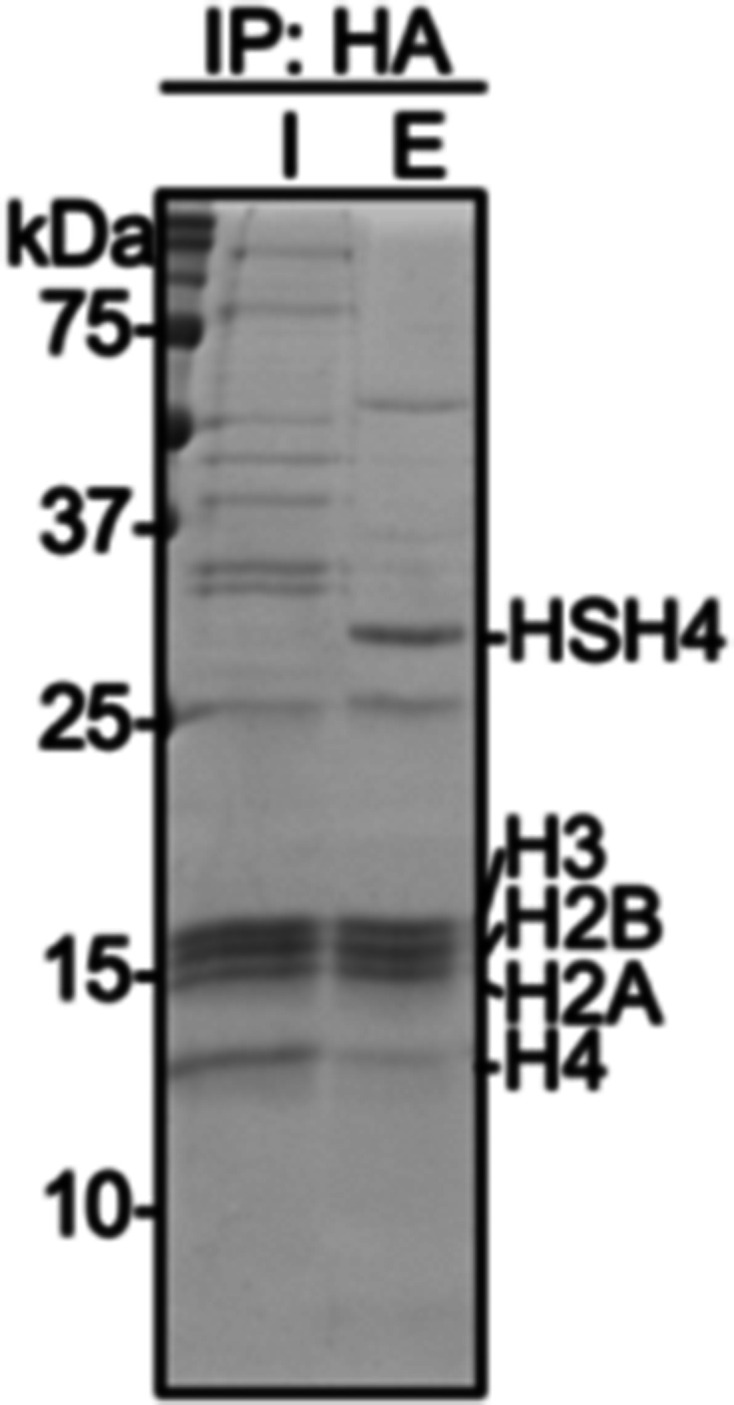
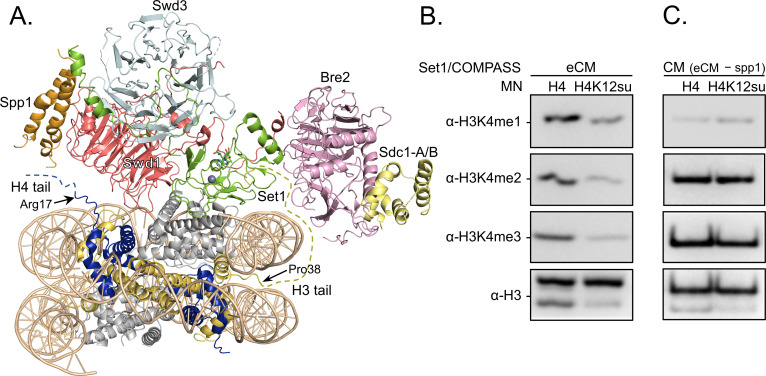
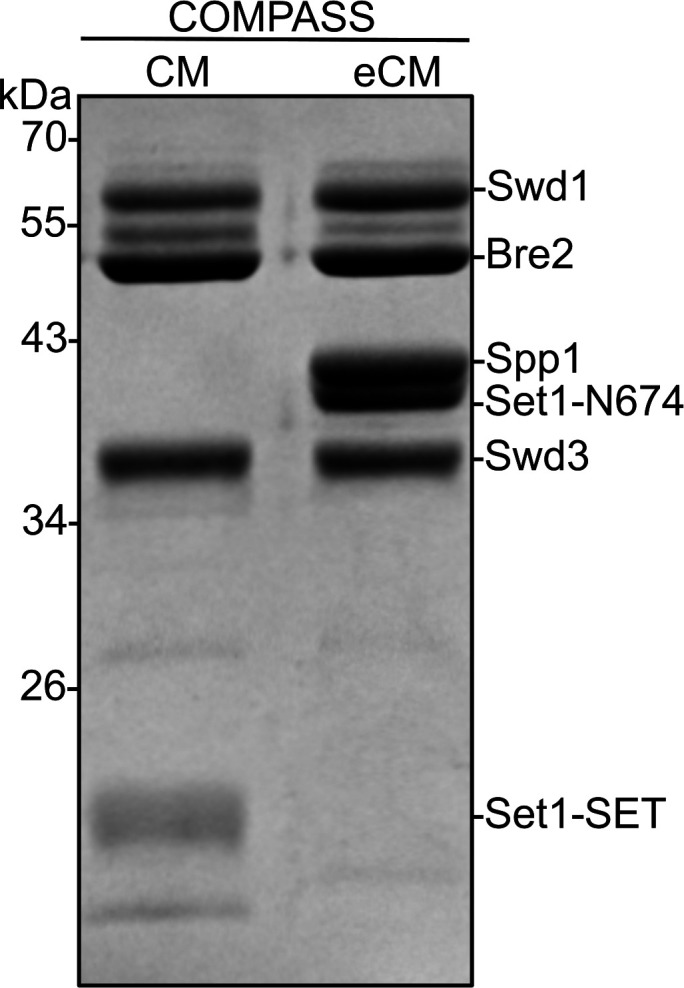
The post-translational modification of histones by the small ubiquitin-like modifier (SUMO) protein has been associated with gene regulation, centromeric localization, and double-strand break repair in eukaryotes. Although sumoylation of histone H4 was specifically associated with gene repression, this could not be proven due to the challenge of site-specifically sumoylating H4 in cells. Biochemical crosstalk between SUMO and other histone modifications, such as H4 acetylation and H3 methylation, that are associated with active genes also remains unclear. We addressed these challenges in mechanistic studies using an H4 chemically modified at Lys12 by SUMO-3 (H4K12su) and incorporated into mononucleosomes and chromatinized plasmids for functional studies. Mononucleosome-based assays revealed that H4K12su inhibits transcription-activating H4 tail acetylation by the histone acetyltransferase p300, as well as transcription-associated H3K4 methylation by the extended catalytic module of the Set1/COMPASS (complex of proteins associated with Set1) histone methyltransferase complex. Activator- and p300-dependent in vitro transcription assays with chromatinized plasmids revealed that H4K12su inhibits both H4 tail acetylation and RNA polymerase II-mediated transcription. Finally, cell-based assays with a SUMO-H4 fusion that mimics H4 tail sumoylation confirmed the negative crosstalk between histone sumoylation and acetylation/methylation. Thus, our studies establish the key role for histone sumoylation in gene silencing and its negative biochemical crosstalk with active transcription-associated marks in human cells.
组蛋白的翻译后修饰由小分子泛素样修饰物(SUMO)蛋白完成,与真核生物中的基因调控、着丝粒定位和双链断裂修复有关。虽然组蛋白 H4 的 SUMO 化与基因抑制特异性相关,但由于在细胞中特异性 SUMO 化 H4 的挑战,这一点无法得到证明。SUMO 与其他组蛋白修饰(如与活性基因相关的 H4 乙酰化和 H3 甲基化)之间的生化串扰也不清楚。我们在使用通过 SUMO-3(H4K12su)化学修饰的 H4(K12 赖氨酸)的机制研究中解决了这些挑战,该修饰被整合到单核小体和染色质化质粒中进行功能研究。基于单核小体的测定表明,H4K12su 抑制转录激活的 H4 尾部乙酰化,该乙酰化由组蛋白乙酰转移酶 p300 完成,也抑制转录相关的 H3K4 甲基化,该甲基化由 Set1/COMPASS(与 Set1 相关的蛋白复合物)组蛋白甲基转移酶复合物的扩展催化模块完成。使用染色质化质粒进行的激活剂和 p300 依赖性体外转录测定显示,H4K12su 抑制 H4 尾部乙酰化和 RNA 聚合酶 II 介导的转录。最后,基于 SUMO-H4 融合的细胞测定证实了组蛋白 SUMO 化与乙酰化/甲基化之间的负生化串扰,该融合模拟了 H4 尾部 SUMO 化。因此,我们的研究确立了组蛋白 SUMO 化在基因沉默中的关键作用及其在人类细胞中与活性转录相关标记的负生化串扰。