Department of Biochemistry and Molecular Biology, University of Kansas Medical Center, Kansas City, Kansas, USA.
Department of Biochemistry and Molecular Biology, University of Kansas Medical Center, Kansas City, Kansas, USA; Molecular, Cellular, and Developmental Biology Program, Division of Biology, Kansas State University, Manhattan, Kansas, USA; Biology & Environmental Health, Missouri Southern State University, Joplin, Missouri, USA.
J Biol Chem. 2022 Jan;298(1):101494. doi: 10.1016/j.jbc.2021.101494. Epub 2021 Dec 14.
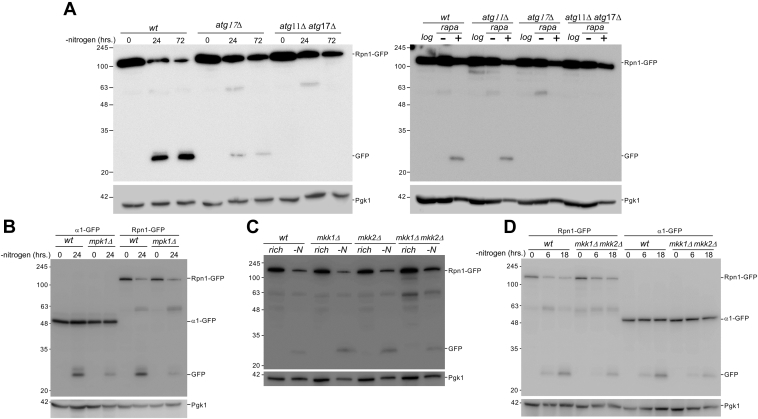
Changing physiological conditions can increase the need for protein degradative capacity in eukaryotic cells. Both the ubiquitin-proteasome system and autophagy contribute to protein degradation. However, these processes can be differently regulated depending on the physiological conditions. Strikingly, proteasomes themselves can be a substrate for autophagy. The signals and molecular mechanisms that govern proteasome autophagy (proteaphagy) are only partly understood. Here, we used immunoblots, native gel analyses, and fluorescent microscopy to understand the regulation of proteaphagy in response to genetic and small molecule-induced perturbations. Our data indicate that chemical inhibition of the master nutrient sensor TORC1 (inhibition of which induces general autophagy) with rapamycin induces a bi-phasic response where proteasome levels are upregulated after an autophagy-dependent reduction. Surprisingly, several conditions that result in inhibited TORC1, such as caffeinine treatment or nitrogen starvation, only induced proteaphagy (i.e., without any proteasome upregulation), suggesting a convergence of signals upstream of proteaphagy under different physiological conditions. Indeed, we found that several conditions that activated general autophagy did not induce proteaphagy, further distinguishing proteaphagy from general autophagy. Consistent with this, we show that Atg11, a selective autophagy receptor, as well as the MAP kinases Mpk1, Mkk1, and Mkk2 all play a role in autophagy of proteasomes, although they are dispensable for general autophagy. Taken together, our data provide new insights into the molecular regulation of proteaphagy by demonstrating that degradation of proteasome complexes is specifically regulated under different autophagy-inducing conditions.
生理条件的改变会增加真核细胞对蛋白质降解能力的需求。泛素-蛋白酶体系统和自噬都有助于蛋白质降解。然而,这些过程的调节可能因生理条件而异。引人注目的是,蛋白酶体本身可以成为自噬的底物。调控蛋白酶体自噬(proteaphagy)的信号和分子机制尚不完全清楚。在这里,我们使用免疫印迹、天然凝胶分析和荧光显微镜来了解响应遗传和小分子诱导的扰动时 proteaphagy 的调节。我们的数据表明,用雷帕霉素抑制主营养素传感器 TORC1(抑制它会诱导普遍自噬)会引起双相反应,其中蛋白酶体水平在自噬依赖性降低后上调。令人惊讶的是,几种导致 TORC1 抑制的条件,如咖啡因处理或氮饥饿,仅诱导 proteaphagy(即没有任何蛋白酶体上调),这表明在不同的生理条件下,proteaphagy 上游信号的会聚。事实上,我们发现,几种激活普遍自噬的条件并没有诱导 proteaphagy,这进一步区分了 proteaphagy 与普遍自噬。与此一致,我们表明,选择性自噬受体 Atg11 以及 MAP 激酶 Mpk1、Mkk1 和 Mkk2 都在蛋白酶体的自噬中起作用,尽管它们对于普遍自噬是可有可无的。总之,我们的数据通过证明蛋白酶体复合物的降解在不同的自噬诱导条件下受到特异性调节,为 proteaphagy 的分子调节提供了新的见解。